
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Хлоропласт. Пластидный или хлоропластный геном.
Хлоропласта - цитоплазматические органеллы растений и водорослей с собственной, хотя и полуавтономной, генетической системой. Синтез соединений, входящих в состав мембран хлоропластов, и процессы, происходящие в ее субкомпартментах, находятся под контролем двух генетических систем: ядра и хлоропласта. Поэтому кратко остановимся на структурных особенностях и специфике функционирования этой клеточной органеллы.
Хлоропласты - внутриклеточные органеллы растений и водорослей, имеющие три типа мембран: высокопроницаемую наружную и менее проницаемую внутреннюю мембрану. Эти две мембраны разделены межмембранным пространством. Внутренняя мембрана окружает центральную область — строму, содержащую ферменты, рибосомы, РНК и ДНК. В хлоропластах имеются также тилакоиды -структуры, напоминающие уплощенные пузырьки, уложенные в стопки (гранулы). В третьей - тилакоидной мембране находятся фотосистемы II и I, электрон-транспортная цепь, связывающая эти системы, и АТР-синтетаза.
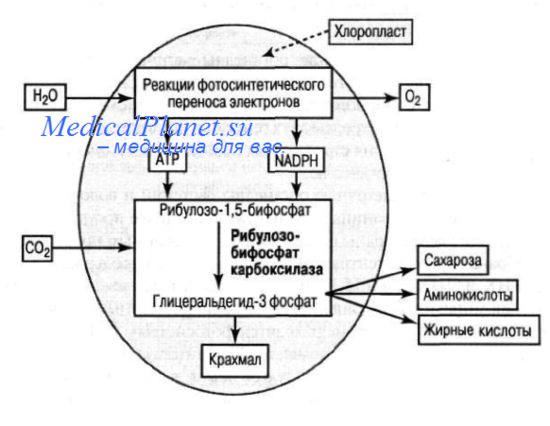
Молекулы хлорофилла, входят, наряду с белками, в состав фотосистем. Фотосистема II катализирует удаление электронов из молекулы воды, а фотосистема 1 ответственна за восстановление NADP+,
В процессе фотосинтеза за счет энергии света образуются молекулы АТР и NADPH, необходимые для превращения С02 в углеводы.
У водорослей и растений число пластид на клетку может превысить два десятка. Районы хлоропласта, в которых находятся молекулы ДНК, по аналогии с прокариотическими клетками, называются иуклеоидами. В клетках растений хлоропластный геном многократно повторен. Подсчитано, что в каждой растительной клетке при наличии более десятка хлоропластов и нескольких молекул ДНК в каждом нуклеоиде, число хлДНК (или геномов) может достигать нескольких тысяч. Увеличение ко-пийностиДНКхлоропластов можно сравнить с полиплоидией -увеличением количества хромосомных наборов в ядре клетки.
ДНК хлоропластов имеет кольцевую форму. Длина молекул хлДНК варьирует от 135 т.п.н. у Euglena gracilis до 200—220 т.п.н. у Pelargonium zonale.
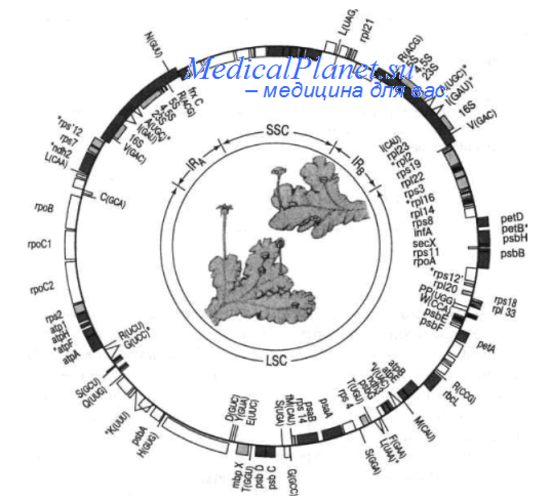
Зеленые растения содержат в хлоропластной ДНК около 100-120 генов. К хлоропластному геному относятся:
• 30 генов тРНК;
• гены pPHK(rRNA)-4.5S;5S; 16S; 23S;
• 20 генов рибосомных белков малой и большой субъединии;
• ген трансляционного фактора IF1;
• часть генов, кодирующих белковые компоненты фотосистемы I и II;
• гены белков электрон-транспортной системы;
• ген субъединицы никотинаминадениннуклеотид (NADН)-дегидрогеназы. Большая часть генов, кодирующих трансляционные факторы, рибосомныс белки, белки фотосистем I и И, белки компонентов электрон-транспортной цепи, связывающей фотосистему II с фотосистемой I, кодируются ядерными генами. Безучастия ядерных генов невозможен синтез белков на рибосомах хлоропластов и осуществление основной функции хлоропластов - фотосинтеза.
При образовании гибридных белков в хлоропластах происходит объединение субъединиц, одни из которых синтезированы на рибосомах цитоплазмы, другие — на рибосомах хлоропласта. Например, фермент рибулозо-1,5- дифосфаткарбоксилаза, участвующий в образовании молекул 3-фосфоглицерата из рибулозо-1,5-дифосфата и С02, состоит из двух субъединиц, одна из которых (rbc L) кодируется хлоропластный геном.
Таким образом, различные элементы белок-синтезирующей системы хлоропластов контролируются либо собственными генами (гены: тРНК, рРНК, фактор IFI, субъединицы никотинаминадениннуклеотид (NADН)-дегидрогеназы), либо ядерными генами (факторы элонгации и терминании трансляции, большинство рибосомных белков), либо находятся под двойным контролем (белковые компоненты фотосистем и белки электрон-транспортной системы).
В хлоропластной ДНК одноклеточной водоросли Chlamydomonas reinhardii имеются гены устойчивости к различным антибиотикам: стрептомицину (sm), эритромицину (еrу), олеандомицину (оlе), неомицину (пеа). Мутации устойчивости к антибиотикам наследуются, как правило, по материнской линии, поскольку в зиготе присутствует только материнская хлДНК. Однако в редких случаях у хламидомонады сохраняется хлоропласт и от второго родителя, тогда зигота становится гетерози-готой. Такие гетерозиготы получили название цнтогеты. При митотическом делении зооспор на стадии четырех нитей, т.е. в удвоенных молекулах хлДНК, между хлоро-пластными генами может происходить рекомбинация. До появления молекулярно-генетических методов картирование хлоропластных генов осуществлялось при изучении частоты рекомбинации между генами и между генами и центромерой.
Наследование пестролистности у растений. Опыты К. Корренса и Э, Баура. Признак белозеленой пестролистности у ночной красавицы наследуется по материнской линии вне зависимости от направления скрещивания. Корренс предположил наличие «смешанных клеток» в пестрых участках ткани, в которых содержатся как зеленые так и бесцветные пластиды. Позднее Реннер и Баур выдвинули гипотезу, согласно которой пластиды (хлоропласты) являются носителями наследственности, и тип окраски передается дочерним пластидам при их делении.
В отличие от ночной красавицы у герани небольшое число пластид передается по мужской линии (двуродительское наследование). Как показали более поздние исследования Тилни—Бассет (1970), наследование окраски листьев находится под ядерным контролем и под контролем пластид женского родителя (мужской родитель оказывает незначительное влияние из-за малого количества пластид в спермиях и большого - в яйцеклетке),
В дальнейших исследованиях было выяснено, что биосинтез хлорофилла, от которого зависит окраска листа, контролируется главным образом ядерными генами. Однако некоторые гены локализованы в пластидной ДНК. Окраска хлорогшастов зависит от фотосинтезируемого пигмента - хлорофилла. Различные формы хлорофилла отличаются по спектрам поглощения. Хлорофиллы а и b обеспечивают зеленую окраску, хлорофилл с — желтую и коричневую.
При действии мутаций, нарушающих биосинтез хлорофилла, в хлоропластах не образуется пигмент. В случае пестролистности материнского растения в яйцеклетке присутствуют пластиды двух типов: нормальные и мутантные, у которых зеленого хлорофилла нет. При делении зиготы пластиды случайно распределяются по дочерним клеткам, в результате листья у нового поколения растений могут быть: зелеными, белыми, либо пестрыми с чередующимися зелеными и белыми участками.
- Читать далее "Митохондрии. Митохондриальный геном."
Оглавление темы "Мутагенез. Внехромосомная наследственность.":1. Закономерности спонтанных мутаций. Течение спонтанного мутационного процесса.
2. Гомологические ряды. Закон гомологических рядов наследственной изменчивости.
3. Антимутагены. Ингибиторы мутагенеза.
4. Выявление мутагенов. Мутагенные факторы окружающей среды.
5. Тестирование на мутагенность. Стратегия тестирования на мутагенность.
6. Генотоксичность. Тест системы на генотоксичность.
7. Стандартизация тест систем на мутагенность. Современные тесты мутагенности.
8. Нехромосомная наследственность. Наследственность не связанная с хромомсомами.
9. Хлоропласт. Пластидный или хлоропластный геном.
10. Митохондрии. Митохондриальный геном.