
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Генотоксичность. Тест системы на генотоксичность.
Известно более 100 различных методов оценки генотоксичности. Однако реально практическое использование имеют не более 20 тест-систем.
Наиболее распространенный метод регистрации влияния ксенобиотиков (мутагенов и канцерогенов) на частоту генных мутаций предложен Брюсом Эймсом в 1975 г. Используемые в качестве тест-объекта His-мутанты Salmonella typhimurium не синтезируют гистидин и выживают на безгистидиновых средах только при возникновении обратной мутации к дикому типу His+. Ревертанты дикого типа образуют колонии на среде без гистидина, что и служит показателем возникновения генных мутаций. Конструирование тестерных штаммов, наиболее чувствительных к действию мутагенов, достигается инактивированием в их клетках системы эксцизионной репарации.
Кроме того, клетки применяемых в тесте Эймса штаммов обладают и иными особенностями, повышающими чувствительность их к мутагенным воздействиям. Многие соединения проявляют мутагенную и канцерогенную активность только при метаболической активации ферментами млекопитающих, (ее проводят в отношении исследуемых соединений in vitro с помощью микросомной фракции печени млекопитающих). В последние годы тесг Эймса был значительно усовершенствован: автоматизирована процедура тестирования, повышена чувствительность к отдельным типам мутагенов.
Принципиально новый подход к оценке генотоксичности - использование трансгенных мышей с интегрированными в геном тестерными генами. По их изменению, выявляемому, например, с помощью рестрикционного анализа, можно оценивать индукцию генных мутаций. Используя трансгенные тест-объекты, можно изучать тканевую и органную специфичность мутагенного действия.
В большинстве случаев у высших организмов мутации отдельных генов не рассматриваются, поскольку они очень редки. Вместо этого оценивают частоту возникновения мутаций в хромосоме в целом.
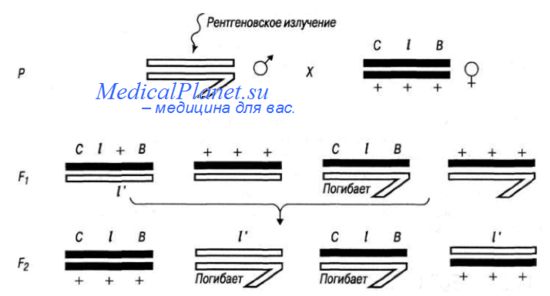
Первый такой метод обнаружения и определения частоты мутаций у дрозофилы -СlВ— применил в (927 г. Герман Меллер. Он разработал систему, позволяющую отличать вновь возникшую мутацию от уже имеющихся в генотипе. Так как наиболее объективно можно учесть частоту рецессивных летальных мутаций, Г. Мёллер ввел в Х-хромосому тестерной линии дрозофилы инверсию Сl, которая играет роль «запирателя» кроссинговера (C) и обладает рецессивным летальным эффектом (l). Кроме того, он маркировал Х-хромосому доминантным геном В (Bar), редуцирующим число фасеток глаза; в результате сферические в норме глаза у самок-гетерозигот приобретают бобовидную форму, а у самцов становятся щелевидными.
Самок тестерной линии СlВ скрещивали с облученными самцами (в качестве мутагена могло выступать и химическое соединение). Из первого поколения выбирали самок СlВ/+ для постановки индивидуальных скрещиваний.
Так как самцы с генотипом ClB/Y погибают независимо от возникновения новой летальной мутации, в случае ее отсутствия расщепление по полу во втором поколении будет 2:1. Отсутствие во втором поколении самцов XY свидетельствует о возникновении в Х-хромосоме летальной мутации. Ее частота выражается отношением числа Х-хромосом, точнее — пробирок с индивидуальными скрещиваниями, где была обнаружена новая летальная мутация, к общему числу пробирок данной выборки.
Следует помнить, что у самок первого поколения между половыми хромосомами иногда может происходить двойной кроссинговер, что приводит к снижению истинной частоты летальных мутаций. В настоящее время метод С1В утратил свое практическое значение.
Вместо него используется предложенный в дальнейшем тем же автором метод Меллер-5. У самок линии-анализатора: обе Х-хромосомы содержат по две инверсии, не связанные с летальным действием: sc8 (редуцированные щетинки) захватывает большую часть Х-хромосомы, 49 - инверсия в средней части Х-хромосомы. Следствием этих инверсий является практически полное исключение перекреста между хромосомами. Дополнительно обе Х-хромосомы самки маркированы геном желтой окраски тела и щетинок yellow (у). Самцы в такой линии жизнеспособны.
Если у взятого для исследования самца дикого типа нет мутации в Х-хромосоме, то, после скрещивания его с самкой из линии-анализатора, во втором поколении мы получим по 2 фенотипических класса самок и самцов. Если же в анализируемой Х-хромосоме исследуемого самца возникла летальная мутация, то во втором поколении все самцы будут принадлежать к одному фенотипическому классу (scsy d49) — желтые с редуцированными щетинками. При этом каждая индивидуальная культура второго поколения, являющаяся потомством одной самки F1 соответствует одной исследованной Х-хромосоме самца из родительского поколения.
Метод Меллер-5, как и тест Эймса, широко применяется для контроля химических соединений, используемых при консервировании пищевых продуктов, изготовлении косметических препаратов и т.п.
Для оценки способности агентов индуцировать хромосомные мутации широко используются цитогенетические методы учета хромосомных аберраций в метафазных клетках пролиферирующих тканей in vitro или in vivo. Недостаток этих методов состоит в том, что они достаточно субъективны (поскольку основаны на микроскопировании), требуют высокой квалификации исследователя и плохо поддаются автоматизации.
В качестве альтернативы был предложен метод учета микроядер (внутриклеточные хроматиновые образования, сформированные из ацентрических фрагментов хромосом и цельных хромосом, отставших в анафазе из-за дефектов веретена деления) в полихроматофильных эритроцитах костного мозга грызунов, который может быть автоматизирован и, кроме того, применен к любой пролиферируюшей ткани, вкчючая гонады.
Для оценки индукции хромосомных мутаций в зародышевых клетках млекопитающих используют учет либо доминантных летальных мутаций, либо наследуемых транслокацнй (последний более специфичен для решении данной задачи).
Очевидно, что максимальное приближение исследователя к оценке генетического риска вследствие действия мутагенов внешней среды возможно только при использовании в качестве тест-систем клеток человека. В таких экспериментах обычно используются лимфоциты периферической крови и, как возможный вариант - клетки костного мозга, эпителия волосяных фолликулов, а также эмбриональные фиб-робласты и сперматозоиды.
- Читать далее "Стандартизация тест систем на мутагенность. Современные тесты мутагенности."
Оглавление темы "Мутагенез. Внехромосомная наследственность.":1. Закономерности спонтанных мутаций. Течение спонтанного мутационного процесса.
2. Гомологические ряды. Закон гомологических рядов наследственной изменчивости.
3. Антимутагены. Ингибиторы мутагенеза.
4. Выявление мутагенов. Мутагенные факторы окружающей среды.
5. Тестирование на мутагенность. Стратегия тестирования на мутагенность.
6. Генотоксичность. Тест системы на генотоксичность.
7. Стандартизация тест систем на мутагенность. Современные тесты мутагенности.
8. Нехромосомная наследственность. Наследственность не связанная с хромомсомами.
9. Хлоропласт. Пластидный или хлоропластный геном.
10. Митохондрии. Митохондриальный геном.