
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Мутации. Теоритические основы мутационной изменчивости.
Несмотря на эффективность систем, поддерживающих идентичность генетического материала при его воспроизведении, консерватизму наследственности всегда сопутствует и противостоит наследственная изменчивость - способность генетического материала претерпевать изменения, наследуемые в потомстве. Наследственная изменчивость организмов состоит из:
• комбинативной изменчивости, обеспечиваемой пере комбинированием генов, хромосом и их сегментов, несущих различные аллели, что выражается в разнообразии организмов-потомков, получивших новые (иные, чем у родителей) комбинации аллелей в результате случайного сочетания при оплодотворении и вследствие кроссинговера, а также
• мутационной изменчивости, являющейся результатом возникающих стойких изменений генов и/или хромосом, которые обусловливают заметные качественные изменения наследственных признаков (подвергаясь отбору, они либо сохраняются в популяциях, либо элиминируются).
Термин «мутация» предложил голландский ботаник Гуго де Фриз в своем классическом труде «Мутационная теория» (1901 —1903 гг.), основные положения которого до сих пор не утратили значения:
• мутации возникают внезапно, дискретно, без переходов;
• они константны в своем проявлении;
• мутации наследуются;
• они могут быть как полезными, так и вредными (добавим, а также - нейтральными);
• выявление мутаций зависит от количества проанализированных особей;
• одни и те же мутации могут возникать повторно, хотя и с низкой частотой. Таким образом, под мутациями подразумеваются дискретные, стабильные изменения наследственного материала, приводящие к изменению фенотипа. Процесс возникновения мутаций называют мутационным, или мутагенезом (последний термин чаще употребляют в отношении индуцированных мутаций). Организм, приобретший какой-либо новый признак и тем самым изменивший свой фенотип в результате мутации, называют мутантом.
Впервые теорию непрерывно идущего в органическом мире мутационного процесса, в результате которого от константных видов «временами отщепляются новые формы», выдвинул СИ. Коржинский — российский академик, директор ботанического сада при Петербургском университете. Книга СИ. Коржинского «Гетерогенезис и эволюция», изданная в России в 1899 г. и переведенная на немецкий язык в 1901 г., стала известна Г. де Фризу в процессе его работы над «Мутационной теорией» и послужила объектом цитирования и обсуждения.
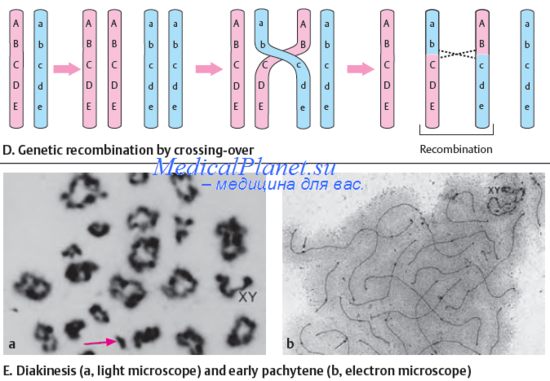
Определение характера и частоты мутаций - тончайший экспериментальный прием, широко используемый генетиками для решения многих фундаментальных проблем. Но особая важность детального изучения мутационного процесса обусловлена тем, что именно мутации служат первоосновой многих наследственных болезней человека. Следовательно, проблема профилактики наследственной патологии хотя бы частично может быть решена только при выяснении механизма становления мутаций.
Охарактеризовать с современных позиций такое сложное, многообразное явление как мутационный процесс чрезвычайно трудно. Сложность этой задачи лучше всего иллюстрирует многообразие подходов к классификации мутаций.
Мутации различают
1) по происхождению:
• спонтанные (возникают самопроизвольно),
• индуцированные (возникают при экспериментальном воздействии на генетический материал);
2) по проявлению в гетерозиготном состоянии:
• доминантные,
• рецессивные;
3) по направлению:
• прямые (переводят состояние дикого типа в качественно иное состояние),
• обратные (иначе — реверсии, возвращают мутантное состояние к дикому типу);
4) по уровню организации изменяемого генетического материала:
• геномные,
• хромосомные,
• генные;
5) по силе проявления аллелей:
• гиперморфные (приводят к усилению действия гена за счет увеличения количества синтезируемого под его контролем продукта),
• гипоморфные (ослабляют действие гена за счет уменьшения количества биохимического продукта, кодируемого аллелем дикого типа),
• неоморфные (кодируют синтез продукта, отличающегося от синтезируемого под контролем аллеля дикого типа, и не взаимодействуют с ним),
• аморфные (инактивируют действие гена),
• антиморфные (действуют противоположно аллелям дикого типа);
6) по влиянию на жизнеспособность и/или плодовитость особей:
• летальные (обусловливают гибель мутанта),
• полулетальные (снижают жизнеспособность, мутанты обычно не доживают до репродуктивного возраста; согласно другому подходу, полулетальные мутации обусловливают гибель половины несущих их особей),
• условно летальные (мутации не проявляются водних —пермиссивных — условиях и лстальны в других - непермиссивных - условиях),
• стерильные (не влияют на жизнеспособность, но резко снижают плодовитость),
• нейтральные (не влияют на жизнеспособность и плодовитость),
• повышающие жизнеспособность и плодовитость особей (жизнеспособность количественно характеризует уровень выживаемости выборки рассматриваемого фенотипического класса по сравнению с другой выборкой в идентичных условиях внешней среды; под плодовитостью подразумевается способность организмов приносить жизнеспособное потомство; часто плодовитость характеризует число потомков одной особи женского пола, родившихся на протяжении ее репродуктивного периода);
7) по характеру регистрируемого проявления:
• морфологические,
• физиологические,
• поведенческие (этологические),
• биохимические и другие (подобное деление мутаций весьма условно: любой признак имеет биохимическую основу, физиологический механизм и морфологическое выражение);
8) по локализации изменяемого генетического материала:
• цитоплазматические (митохондриальные, пластидные),
• ядерные;
9) по месту возникновения и характеру наследования:
• генеративные (они возникают в клетках полового зачатка и в половых клетках и передаются по наследству. При этом мутация, появившаяся на стадии яйцеклетки или сперматозоида, останется единичной, а мутация, возникшая на ранней стадии оогенеза или сперматогенеза, размножится в количестве, пропорциональном числу прошедших клеточных делений, при этом часть зрелых половых клеток будет нести мутантный аллель, а у другой части генотип останется неизмененным);
• соматические (они возникают в соматических клетках и либо приводят к появлению мозаиков/химер у организмов, размножающихся исключительно половым путем, либо наследуются у организмов, имеющих бесполое размножение. Если из мутировавшей соматической клетки растения развивается почка, а из нее — побег, то он будет нести мутантный признак и в перспективе может дать начало новому виду, а в случае селекции — новому сорту).
- Читать далее "Геномные мутации. Гаплоидия. Полиплоидия."
Оглавление темы "Генные и хромосомные мутации.":1. Регуляция транскрипции у прокариот. Негативная и позитивная регуляция генной активности.
2. Специфическая регуляция генной активности. Методы регуляции генной активности.
3. Неспецифическая регуляция генной активности. Компенсация дозы генов у дрозофилы.
4. Компенсация дозы генов у млекопитающих. Современная теория инактивации Х-хромосомы.
5. Регуляция генной активности на уровне репликации. Трансляционная и посттрансляционная регуляция генной активности.
6. Мутации. Теоритические основы мутационной изменчивости.
7. Геномные мутации. Гаплоидия. Полиплоидия.
8. Анеуплоидия. Нуллисомия. Моносомия. Полисемия.
9. Хромосомные мутации. Делеции. Дупликации.
10. Инверсии хромосом. Транслокации хромосом.