
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Специфическая регуляция генной активности. Методы регуляции генной активности.
Промоторы. Регуляция экспрессии генов эукариот основана на тех же принципах, что и у прокариот, хотя существуют и различия. Во-первых, как уже упоминалось, у эукариот нет оперонов, и каждый ген представляет собой независимую транскрипционную единицу. Промоторы эукариот несколько отличаются от прокариотических по структуре. Функционально наиболее важные последовательности в промоторе эукариотического гена находятся в положениях «-25" (бокс Хогнесса) и «-75». Есть и другие участки, более удаленные от стартовой точки транскрипции.
Другую структуру имеют промоторы генов транспортных, рибосомных РНК и 5SPHK. Прежде всего, эти промоторы внутренние, поэтому перед номерами их нуклеотидов ставится знак «+». Промотор гена тРНК представлен на рисунке. В промоторе генатРНК имеются А- и В-боксы. Установлены их канонические последовательности. При уменьшении расстояния между боксами транскрипция снижается или полностью прекращается. Замена всего одного нуклеотида в В-боксе в положении 56 CG —> GC в TG петле искажает правильную структуру тРН К. Регуляторная область гена 5S имеет сходную структуру. Она расположена между +50- и + 90-нуклеотидами и также имеет А-и С-боксы.
Энхансеры. Среди регуляторных элементов эукариот выделяют энхансеры, которые обычно располагаются достаточно далеко от регулируемого гена. Предполагается, что сближение энхансера и промотора достигается в результате образования между ними петли ДН К, при этом белки-активаторы, «узнающие» энхансер, могут непосредственно взаимодействовать с транскрипционным комплексом.
Энхансеры - усилители транскрипции - обладают следующими свойствами:
• могут находиться как в 5', так и в 3'-областях, а также в интронах и даже на значительном расстоянии от промоторов;
• активируют гены независимо от ориентации;
• один энхансер может активировать различные гены;
• действие их может быть ткане- и видоспецифичным;
• энхансеры доступны действию различных белков, в том числе и гормонов.
Сайленсеры - ослабители транскрипции — являются негативными элементами по отношению к транскрипции. Они так же, как и энхансеры, функционируют в цис-положении и могут оказывать свое действие на большом расстоянии от гена и при разной ориентации по отношению к нему.
Транскрипционные факторы. Многоклеточные эукариоты состоят из различных типов клеток, разнообразие которых обусловлено дифференциальной экспрессией генов, определяющих образование тканеспецифических белков. Один из механизмов, лежащих в основе дифференциальной экспрессии, - регуляция процесса транскрипции.
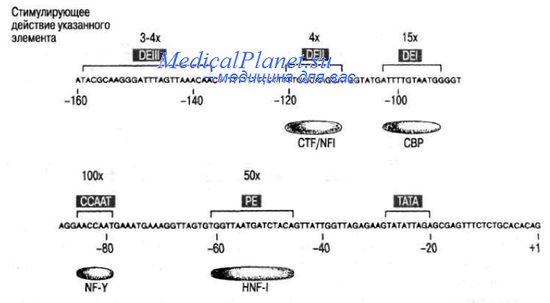
Так, изучено влияние регуляторных белков на активность промотора альбуминового гена AL В млекопитающих, функционирующего в клетках печени. Левее стартовой точки расположены: последовательность промотора TATA (определяющего начало транскрипции, но не частоту инициации этого процесса) и последовательность ССААТ (влияющая на эффективность транскрипции). Тканевая специфичность определяется взаимодействием специфических факторов транскрипции со вспомогательными последовательностями промотора РЕ, DEI, DEI), и DEIII. Последовательность РЕ у мыши (из 13 нуклеотидов) служит сайтом связывания для тканеспецифического белка печени - HNFI. С последовательностью DEI взаимодействует белок СВР, а с DEII - NFI. Эти белки не обладают специфичностью, их роль - активация процесса транскрипции. Пока не ясно, каким образом взаимодействуют специфические и неспецифические белки в регуляторной области гена ALB.
Установлены сайты связывания регуляторных белков в промоторе гена Р-интерферона IFN. Промотор этого гена находится между парами оснований -204 и +1. Левее ТАТА-бокса расположены последовательности пяти элементов, две из которых (NRDI и NRD2) ответственны за негативную регуляцию, а три (PRDI, PRD1I и PRDIII) способствуют активации транскрипции при присоединении к ним соответствующих белков. Механизм смены этих процессов пока не изучен.
Факторы транскрипции и ядерный матрикс. Ядерный матрикс был открыт в 50-е гады прошлого века И.Б. Збарским. Эта структура состоит из нерастворимых белков. Именно на нем происходит процесс транскрипции и созревания пре-мРНК. Показано, что факторы транскрипции, соединяясь с ядерным матриксом, обеспечивают правильное пространственное расположение промоторньгх и энхансерных участков генов. Они взаимодействуют с ДНК в участках связывания с матриксом и другими белками ядерного матрикса, которых выявлено не менее 50. К ним относятся белки, связывающиеся с ДНК посредством «цинковых пальцев» (SP-l, MyTl, ZNF74, ATRX); белки, связывающиеся с ДНК с помощью «лейциновой застежки-молнии» (С/ЕВР, ATF, YY- i); рецепторы стероидных гормонов и ассоциированные с ними факторы; белки с HMG-доменом (HMGl, HMG2, HMG14, HMG17, AVL-1B, NMP2) и многие другие.
Часто транскрипционные регуляторные факторы имеют специфическую структуру, например, структуру типа "цинковых пальцев". Она характеризуется аминокислотными петлями, имеющими в основании два цистеина в одной части петли и два гистидина - в другой, связанные ионом цинка. Наряду с такими ферментами как РН К-полимеразы и топоизомеразы, в клетках эукариот присутствуют различные регуляторные белки, взаимодействующие с промоторами, энхансерами и сайленсерами. Таким белком является PI, взаимодействующий с CCGCCC-блоком и ядерный фактор, связывающийся с СААТ-блоком. Нуклеосомьг в активном хроматине соединены с белками HMG14H HMG17.
В научной литературе обсуждаются различные гипотезы, обьясняющие, как РН К-полимераза может транскрибировать хроматин, В соответствии с одной из них, нуклеосома разворачивается, половинки ее сердцевины разъединяются, РНК-полимераза транскрибирует ДНК, а затем половинки нуклеосомы опять складываются. Согласно другой модели, РНК-полимераза транскрибирует витки ДНК, которые нуклеосома сбрасывает по очереди: сначала один виток, а затем - другой.
Метилирование оснований ДНК. Значительную роль в регуляции экспрессии генов у эукариот может играть метилирование ДНК (обычно по 5-му углероду цитозина). Неактивные гены содержат относительно много метильных групп. Состояние повышенного метилирования может стабильно поддерживаться в течение многих поколений клеток. Для этого существует специальный механизм, обеспечивающий присоединение метильных групп в местах, аналогичных тем, где уже произошло метилирование в другой цепи. Механизм действия метильных групп заключается в том, что они нарушают взаимодействия ДНК-белок. Выступая в большую бороздку ДНК, метильные группы препятствуют связыванию транскрипционных факторов. Кроме того, метилированные районы ДНК могут взаимодействовать с транскрипционными репрессорами типа МеСР2, являющегося составной частью белкового регуляторного комплекса.
Импринтинг. Феномен импринтинга демонстрирует роль метилирования в регуляции экспрессии генов. Суть этого феномена заключается в том, что аллели, унаследованные от отца и матери, экспрессируются по-разному. Согласно одной из гипотез, это связано с различным метилированием аллелей при образовании половых клеток. Подробно болезни импринтинга рассмотрены в части II. Медицинская генетика.
Другой пример влияния метилирования на активность генов — лайонизация одной из Х-хромосом у женщин. Гены инактивированной (лайонизированной) хромосомы почти все метилированы.
- Читать далее "Неспецифическая регуляция генной активности. Компенсация дозы генов у дрозофилы."
Оглавление темы "Генные и хромосомные мутации.":1. Регуляция транскрипции у прокариот. Негативная и позитивная регуляция генной активности.
2. Специфическая регуляция генной активности. Методы регуляции генной активности.
3. Неспецифическая регуляция генной активности. Компенсация дозы генов у дрозофилы.
4. Компенсация дозы генов у млекопитающих. Современная теория инактивации Х-хромосомы.
5. Регуляция генной активности на уровне репликации. Трансляционная и посттрансляционная регуляция генной активности.
6. Мутации. Теоритические основы мутационной изменчивости.
7. Геномные мутации. Гаплоидия. Полиплоидия.
8. Анеуплоидия. Нуллисомия. Моносомия. Полисемия.
9. Хромосомные мутации. Делеции. Дупликации.
10. Инверсии хромосом. Транслокации хромосом.