
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Сцепленное наследование. Полное сцепление. Неполное сцепление. Кроссинговер.
Под сцеплением генов генетики понимают совместное наследование генов, локализованных в одной хромосоме. Независимо (как это было в опытах Г. Менделя) неаллельные гены ведут себя только в том случае, если они находятся в негомолотчных хромосомах. Тогда их поведение объясняется независимым расхождением и свободной комбинаторикой негомологичных хромосом в мейозе.
Впервые сцепленное наследование признаков в дигибридном скрещивании обнаружили У. Бэтсон и Р. Пеннет удушистого горошка Lathynts odoratus при изучении наследования окраски цветка и формы пыльцевых зерен. Классическими же опытами по анализу сцепленного наследования были эксперименты Т. Моргана, исследовавшего закономерности наследования мутаций black (b — черное тело) и vestigial (vg — редуцированные крылья) у D. melanogasler. Аллели дикого типа определяют серую окраску тела (b+) и нормальные крылья (vg+) и доминируют над мутантными аллелями. При скрещивании самок с черным телом и нормальными крыльями (b vg+/b vg+) с серыми самцами, имеющими редуцированные крылья (b+ vg/b+ vg), все потомство F, имело признаки дикого типа,

Результаты анализирующего скрещивания потомков F, оказываются разными. В зависимости от пола гибридов обнаруживается или полное сцепление генов, или неполное.
Если для анализирующего скрещивания взять дигетерозиготного самца F, и самку, гомозиготную по рецессивным аллелям (b vg/b vg), то получится только два класса потомков; 50% мух серых с редуцированными крыльями и 50% - черных с нормальными крыльями.
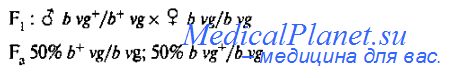
Это означает, что у дигетерозиготных самцов bvg+/b+vg образуется только два типа гамет (bvg+ и b+vg). Если бы гены b и vg наследовались независимо, то дигетеро-зиготные самцы F1 дали бы 4 типа гамет (b vg, b vg+, b+ vg, b+ vg+) и, соответственно в потомстве анализирующего скрещивания были бы представлены 4 фенотипических класса в соотношении 1:1:1:1. Морган в своих опытах показал, что гены могут наследоваться сцепленно, что приводит к появлению в потомстве только родительских комбинаций генов (b vg+ и b+vg). У самцов дрозофилы выявляется полное сцепление генов.
При реципрокном направлении скрещивания (самки - дигетерозиготы, самцы - гомозиготы по рецессивным аллелям обоих генов) результаты анализирующего скрещивания иные:
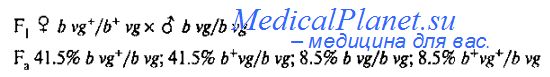
Сочетания признаков исходных родительских линий (Р-поколения), сохранившиеся у потомков анализирующего скрещивания, являются нерекомбинантными: b vg+ (черное тело, нормальные крылья) и b+vg (серое тело, редуцированные крылья), Видно, что родительские сочетания образовались в 83% случаев. Новые, рекомбинантные, сочетания b+vg+ и b vg составляют 17% потомков. Таким образом, в данном случае наблюдается неполное сцепление генов.
Полное сцепление у дрозофилы наблюдается только к том случае, когда для анализирующего скрещивания берут дигибридных самцов, если же скрещивают дигибридных самок с гомозиготными по рецессивным аллелям самцами, то сцепление никогда не бывает полным.
Кроссинговер
Опираясь на наблюдения Ф. Янсенса (1909), обнаружившего в мейозе хиазмы, Т. Морган постулировал, что гены b и vg локализованы в одной хромосоме, и в потомстве анализирующего скрещивания особи b vg/b vg и b+ vg+/b vg появились в результате перекреста гомологичных хромосом и обмена участками между генами b и vg. Процесс перекреста был назван кроссинговером. Отсутствие рекомбинантных потомков (и, следовательно, хромосом) в потомстве самцов объясняется тем, что у дрозофилы при сперматогенезе кроссинговер не происходит Таким образом, описанный эксперимент Моргана и его сотрудников позволил сделать вывод о соответствии групп сцепления и хромосом и явился генетическим доказательством кроссинговера. Кроссинговер проявляется как неполное сцепление. Вскоре феномен кроссинговера был обнаружен и у других организмов - кукурузы, львиного зева, мышей и т.д.
Генетические и цитологические исследования, проведенные на большом числе видов животных и растений, позволили сформулировать следующие положения о механизме кроссинговера. Кроссинговер между несестринскими хроматидами происходит в мейозе на стадии четырех нитей. В каждой точке обмена происходит разрыв и воссоединение только двух из четырех хроматид. На рисунке видно, что из четырех продуктов мейоза две хроматиды являются рекомбинантными, две другие сохраняют родительские комбинации генов. На участке между генами А и В может происходить два и больше обменов, если гены достаточно далеко отстоят друг от друга. Во множественных обменах могут участвовать две, три и четыре хроматиды.
Если гены расположены так далеко друг от друга, что вероятность обмена между ними равна 100%, то 50% гамет будет нести хромосомы с родительской комбинацией генов (некроссоверные), а 50% будут кроссоверными, с новой комбинацией генов, и в потомстве будет обнаруживаться соотношение 1:1:1:1. Таким образом, обнаруживаемая частота рекомбинации между двумя генами не превышает 50%, даже если между генами происходят и множественные обмены.
- Читать далее "Расстояние между генами. Двойной кроссинговер. Интерференция генов."
Оглавление темы "Генотип и кроссинговер.":1. Генотип и фенотип. История изучения генотипа и фенотипа.
2. Вариация наследственных признаков. Фенотип и развитие организма.
3. Гетерогенные группы. Генокопии и фенокопии. Неоморфизм. Гипоморфизм. Антиморфизм.
4. Пенетрантностъ признака. Экспрессивность признака.
5. Неустойчивая и условная доминантность в генетике. Множественные аллели признака.
6. Сцепленное наследование. Полное сцепление. Неполное сцепление. Кроссинговер.
7. Расстояние между генами. Двойной кроссинговер. Интерференция генов.
8. Генетические карты. Картирование генов.
9. Сравнение карт. Неравный кроссинговер. Соматический кроссинговер.
10. Частота кроссинговера. Факторы влияющие на кроссинговер.