
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Генетика и онтогенез. История изучения генетики и онтогенеза.
Онтогенез многоклеточных (Metabiota) представляет собой удивительный по сложности и стройности комплекс ростовых и формообразовательных процессов, в результате которых развившийся из зиготы организм наследует соответствующие видоспецифические и индивидуальные черты организации, физиологии, а в случае человека - и психики.
При этом различают ростовые процессы на клеточном уровне (увеличение количества клеток в результате делений, увеличение клеточного объема) и морфогенез (формообразование), включающий как рост, так и локальную гибель клеток; изменение их формы; движения и изгибы клеточных пластов и многое другое. То, что индивидуальное развитие организма находится под генетическим контролем, доказывается фактом существования наследственной изменчивости по всем рассматриваемым процессам. Так, выявлены мутации генов, контролирующих скорость первых делений дробления, обнаружены онкогены, регулирующие ростовые процессы, известно множество мутаций генов, ответственных за становление морфологических признаков организма в ходе онтогенеза.
Современные исследования онтогенеза многоклеточных на молекулярном уровне весьма обширны, но на вопрос, как из двух клеток (яйцеклетки и спермия) развивается организм, представленный практически двумя сотнями гистотипов клеток, точного ответа пока нет. Как говорил Н .В. Тимофеев-Ресовский: "...почему в процессе развития Metabiota в должное время в должном месте происходит должное?". Это был и есть главный вопрос теории онтогенеза.
Первые попытки ответить на него восходят еще к средним векам, когда оформились две основные теории:
1. Преформизм. Сторонники этой теории утверждали, что развития как такового нет, и каждый индивид представляет собой, грубо говоря, матрешку, т.е. его развитие - это развертывание и рост уже содержащихся в яйцеклетке иди спермин специфических микроструктур. Согласно А. Гадлеру и Ш. Боннэ, в каждой яйцеклетке содержится полностью сформированный зародыш, и каждый такой зародыш имеет яичник с яйцеклетками, в которых в свою очередь содержатся еще меньшие зародыши. По подсчетам А. Галлера, в яичнике Евы должно было содержаться около 200 млрд. зародышей.
2. Эпигенез. Сторонники этой теории полагали, что внутренний механизм развития отсутствует, в исходной клетке ничего нет, все появляется в процессе взаимодействия со средой. Они настаивали на том, что организм в целом и отдельные органы в частности по мере развития не просто увеличиваются в размерах, но и усложняются, К.М. Бэру принадлежит детальное описание развития куриного эмбриона как строго упорядоченного процесса последовательных изменений от яйца к зародышу И далее к зрелой особи.
Хотя ясно, что теория эпигенеза во многих отношениях более корректна, чем взгляды преформистов, но даже в 70-х годах XX века некоторые исследователи отрицали генетическую регуляцию индивидуального развития, полагая, что геном только сопутствует развитию, а не определяет его. Тем не менее, и эпигенез, и преформизм вкупе имеют рациональное зерно: развитие многоклеточных может быть представлено как преформированный эпигенез, или эпигенетическая преформация.
Зигота действительно не содержит никакого предварительно сформированного зародыша. От родителей потомству достается ряд «указаний» в виде заключенной в ДНК генетической информации, которая во взаимодействии с окружением направляет ход развития организма.
Содержат ли все соматические клетки одинаковый набор генов? Или эти наборы различаются? Вильгельм Ру, один из создателей ядерной теории наследственности, предположил, что ядра, возникающие при дроблении зиготы, разнокачественны, а дифференцировка клеточных функций представляет собой результат расхождения в разные клеточные ядра качественно различного наследственного материала. Согласно его теории, сформулированной в современных терминах, каждая клетка содержит в своем ядре только те гены, которые необходимы для программирования характерных для нее функций. Специализация в развитии будет, таким образом, результатом постепенного формирования мозаики клеток, содержащих различные части генома.
Однако его современник Г. Дриш, проверяя в эксперименте гипотезу о том, что у Metabiota ядро каждой живой клетки содержит тот же самый геном, что и ядро зиготы, показал, что перемещение ядер между клетками эктодермы и мезодермы дробящегося зародыша не нарушает его нормального развития. Зачаток регенерирующего хвоста тритона может быть пересажен в область конечности и превратится в ногу, а не в хвост. Следовательно, дробление и последующая дифференцировка не сопровождаются утерей или необратимыми изменениями ядерного материала.
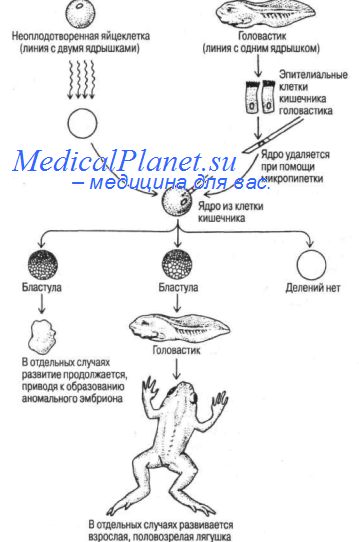
Тотипотентность дифференцированных соматических клеток, т.е. их способность обеспечивать полное развитие организма свойственна и животным, и растениям. Одно из наиболее прямых доказательств этого факта — опыты английского генетика Дж. Гердона по трансплантации ядра из дифференцированной соматической клетки в безъядерную зиготу. Используя африканскую шпорцевую лягушку Xenopus laevis, он получил взрослых особей на основе генетической информации ядра соматической клетки. Путем облучения большими дозами УФ из неоплодотворенных яиц функционально удалялось ядро; затем в каждое из энуклеированных яиц вводили дифференцированное ядро из клетки кишечного эпителия головастика, и таким способом инициировали развитие. В ряде случаев из подобных яиц развивались головастики, а затем и взрослые лягушки.
Для доказательства того, что в развитии участвовало именно трансплантированное, а не собственное ядро яйцеклетки (выжившее при УФ-облучении), применяли генетическое маркирование. Для выделения яйцеклеток использовали линию, характеризующуюся наличием в ядре двух ядрышек - по одному на каждый ядрышковый организатор двух гомологичных хромосом. Ядро соматической клетки было получено от особи, гетерозиготной по делеции ядрышкового организатора и потому имеющей в ядре только одно ядрышко. Все лягушки, развившиеся в результате пересадки ядер, имели по одному ядрышку. Несколько взрослых лягушек из их числа достигли половозрелости, и оказались вполне фертильными, как и нормальные Xenopus.
Эти эксперименты показали, что дифференцировка клеток в онтогенезе не обязательно сопровождается необратимой инактивацией генетического материала ядра, ядро соматической клетки с репрессированной большей частью генома может претерпеть дерепрессию, если его трансплантировать в ооплазму оплодотворенной яйцеклетки, способной к реализации полного цикла развития.
В экспериментах нескольких типов было доказано, что вся информация, необходимая для нормального развития, присутствует в ядре дифференцированной клетки и может быть вновь активирована и использована для повторения процесса развития. Таким образом, проблема генетического контроля индивидуального развития перешла в рамки проблемы дифференциальной экспрессии генов, которая может происходить на уровне любого известного матричного процесса. Концепция дифференциальной активности генов была выдвинута в начале 30-х годов XX в. Т. Морганом. Согласно этой концепции во всех клетках организма имеется одинаковый набор генов, но функционируют они в различно дифференцированных клетках по-разному. В настоящее время дифференциальная активность генов в клетках и тканях разной специализации и на разных этапах онтогенеза - это экспериментально установленный факт универсального значения: в каждом данном типе клеток одновременно экспрессируется лишь небольшая доля генов, и спектры экспрессирующихся генов в клетках разных типов весьма специфичны.
Очевидно, различные регуляторные сигналы, обеспечивающие «включение» и выключение генов, представляют собой важную составную часть механизма дифференцировки клеток, которая происходит на основе детерминации определенного клеточного типа. Хотя накопленные знания все еще не позволяют полностью представить себе картину онтогенеза, некоторые ее фрагменты и связи между ними уже ясны.
Видео урок истории отечественной генетики - И.И. Вавилов, Т.Д. Лысенко
- Читать далее "Этапы онтогенеза. Детерминация в онтогенезе."
Оглавление темы "Онтогенез и генетика.":1. Генетический контроль. Мужская стерильность растений.
2. Геном митохондрий человека. Митохондриальный геном человека.
3. Происхождение митохондрий. Откуда появились митохондрии?
4. Материнский эффект цитоплазмы. Влияние цитоплазмы на генетический материал.
5. Генетика и онтогенез. История изучения генетики и онтогенеза.
6. Этапы онтогенеза. Детерминация в онтогенезе.
7. Ооплазматическая сегрегация. Созревание ооцита.
8. Генетический контроль сегментации. Гены и процесс сегментации организма.
9. Гомеозис. Гомеозисные гены.
10. Гомеобоксы у человека. Наследственные болезни.