
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Структура и анатомия волос. Типы кератина в волосяном фолликуле
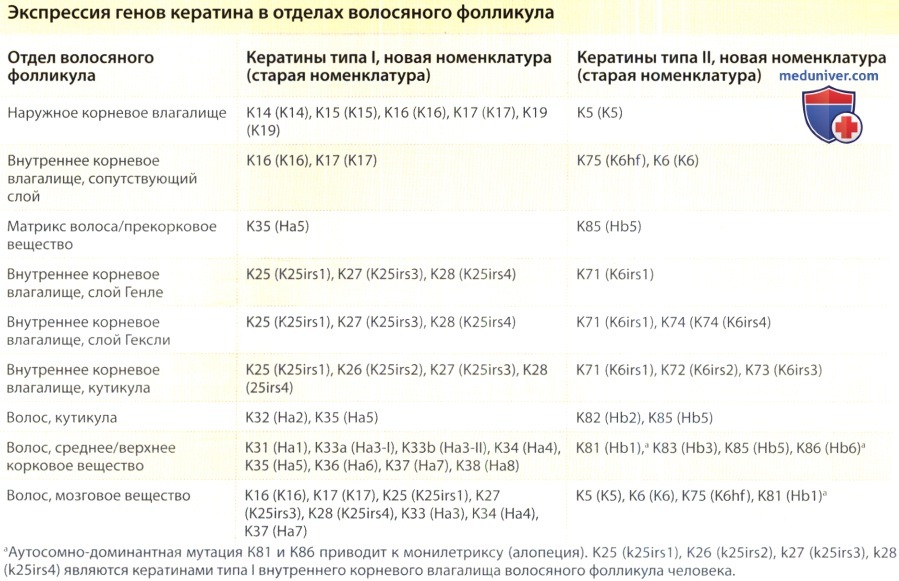
Верхний фолликул состоит из воронки и перешейка, а в нижнем выделяются супрабульбарный (надлуковичный) и бульбарный участки. Верхний фолликул является постоянным, а нижний регенерируется с каждым циклом роста волоса. Основные, составляющие волос части, начиная от самой наружной и оканчивая самой глубокой внутренней, включают соединительнотканное влагалище, наружное корневое влагалище, внутреннее корневое влагалище, кутикулу, кору и сердцевину (мозговое вещество) волосяного стрежня, причем для каждой из этих частей характерна экспрессия специфических для волосяного фолликула кератинов.
а) Наружное корневое влагалище. Наружное корневое влагалище является продолжением эпидермиса в волосяной воронке и простирается вниз до луковицы. Клетки наружного корневого влагалища на протяжении фолликула существенно изменяются. Наружное корневое влагалище в волосяной воронке напоминает эпидермис и образует зернистый слой в процессе кератинизации. В перешейке клетки наружного корневого влагалища подвергаются кератинизации по трихилеммальному типу с отсутствием зернистого слоя. Трихилеммальная кератинизация происходит там, где внутреннее корневое влагалище начинает слущиваться.
Экспрессия десмоглеина здесь также значительно изменяется и трихилеммальные или волосяные кисты сохраняют эти характеристики. Кератиноциты наружного корневого влагалища образуют вздутие (сумку) у основания перешейка. Эти клетки обычно имеют более высокое соотношение ядра и цитоплазмы по сравнению с клетками в других участках фолликула. По мере движения вниз клетки наружного корневого влагалища становятся намного крупнее и в надлуко-вичной части фолликула содержат большое количество гликогена. В луковице наружное корневое влагалище состоит только из одного уплощенного слоя клеток, который можно проследить до основания фолликула.
б) Внутреннее корневое влагалище. Внутреннее корневое влагалище простирается от основания луковицы до перешейка и состоит из четырех слоев (начиная от самого наружного и кончая самым глубоким внутренним): сопутствующего слоя, слоя Генле, слоя Гекели и кутикулы внутреннего волосяного фолликула. Сопутствующий слой считался самым глубоким внутренним слоем наружного корневого влагалища, но последние данные указывают на то, что он больше похож на внутреннее, чем на наружное корневое влагалище. Сопутствующий слой примыкает к слою Генле и идет вверх вместе с остальными слоями внутреннего корневого влагалища, создавая таким образом плоскость скольжения между неподвижным наружным корневым влагалищем и внутренним корневым влагалищем.
В некоторых фолликулах сопутствующий слой выражен сильнее, чем в других слоях (например, в области роста бороды). Клетки сопутствующего слоя плоские, в отличие от кубоидных клеток наружного корневого влагалища, и экспрессируют цитокератин типа II K6hf.75 Слой Генле состоит из одного слоя клеток. Это первый слой, в котором развиваются гранулы кератогиалина, и происходит кератинизация. Слой Гекели состоит из двух или четырех слоев клеток, его кератинизация происходит над слоем Генле в участке, известном как «полоса Адамсона» (Adamson’s fringe). Некоторые клетки в слое Гекели проникают через слой Генле и примыкают непосредственно к сопутствующему слою. Эти клетки называются «крылатыми клетками».
Клетки кутикулы внутреннего корневого влагалища частично накладываются друг на друга, создавая картину «черепичной крыши» и четко переплетаются с клетками кутикулы волосяного стержня. Эта связь между двумя кутикулами плотно удерживает волосяной стержень в фолликуле. Внутреннее корневое влагалище, состоящее из твердых кератинов и связанных с ними белков, придает форму волосу и направление клеткам волосяного стержня по мере их появления. Решающую роль в дифференцировке клеток и клеточных линий внутреннего корневого влагалища играет фактор транскрипции GATA-3. У мышей с отсутствием этого гена внутреннее корневое влагалище не формируется.
в) Волосяной стержень. Волосяной стержень (как и внутренне корневое влагалище) происходит из быстро пролиферирующих матриксных кератиноцитов луковицы, скорость пролиферации которых одна из самых высоких в организме человека. Клетки будущего волосяного стержня располагаются на верхушке дермального сосочка и впоследствии образуют мозговое и корковое вещество, а также кутикулу волосяного стержня. Сразу же над матриксными клетками, в предкератогенной зоне, клетки волосяного стержня начинают экспрессировать специфические для волосяного стержня кератины.
Дифференцировка клеток волосяного стержня в этой зоне зависит от транскрипционного фактора Lef-1. Участки связывания Lef-1 присутствуют в большинстве генов кератина. Рецептор BMP типа 1А также важен для дифференцировки матриксных клеток в волосяной стержень, поскольку потеря этого рецептора препятствует дифференцировке волосяного стержня.
Кутикула волосяного стержня покрывает волос, и ее целостность и свойства существенно влияют на внешний вид волос. Как только волос появляется на коже волосистой части кожи головы, кутикула испытывает воздействие внешней среды и часто полностью утрачивается на дистальных участках длинных волос. Корковое вещество внутри кутикулы составляет основную массу стержня и содержит меланин. Корковое вещество расположено в форме длинных шнуровидных структур, называемых макрофибриллами. Они, в свою очередь, включают микрофибриллы, которые состоят их промежуточных филаментов. Мозговое вещество расположено в центре более крупных волос, а специфические кератины, которые экспрессированы в этом слое клеток, находятся под контролем андрогенов.
г) Дермальный сосочек. Дермальный сосочек является сердцевиной, происходящей из мезенхимы ткани, которая покрыта оболочкой матриксного эпителия. Он состоит из фибробластов, пучков коллагена, мукополисахаридной стромы, нервных волокон и одиночной капиллярной петли. Дермальный сосочек является продолжением соединительнотканного перифолликулярного влагалища (дермального влагалища), которым окружен нижний фолликул.
Эксперименты по рекомбинации тканей показали, что дермальный сосочек обладает мощными индуктивными свойствами, включая способность индуцировать формирование волосяного фолликула при трансплантации в область не имеющего волосяного покрова эпидермиса на подушечках лап у мышей. Это свидетельствует о том, что тканевая паттернизация, сформировавшаяся во время эмбрионального периода может, при соответствующих условиях, изменяться. В волосяном фолликуле человека объем дермального сосочка коррелирует с количеством матриксных клеток и возникающим в результате их функции размером волосяного стержня. У мышей размер волосяной луковицы и диаметр волос во многом зависят от пролиферативной активности матриксных кератиноцитов.
Многие растворимые факторы роста происходят из дермального сосочка, который паракринно воздействует на расположенные сверху матриксные клетки эпителия. В частности, дермальным сосочком в фазе анагена генерируется фактор роста кератиноцитов (KFG), а его рецептор, рецептор фактора роста фибробластов 2 (FGFR2), локализуется преимущественно в матриксных кератиноцитах. Инъекции KFG голым мышам вызывают усиленный рост волос в участках инъекции, что указывает на вероятную необходимость KFG для роста волос и волосяного цикла. Удивительно, однако, что у мышей, нокаутных по KFG, развиваются морфологически нормальные волосяные фолликулы, которые производят «шершавые» или «жирные» волосы. Следовательно, воздействие KGF на морфогенез волосяного фолликула и цикл роста волос может замещаться действием других факторов роста с избыточными функциями.
д) Иннервация волосяного фолликула. Миелиновые сенсорные нервные волокна идут параллельно волосяным фолликулам, образуя сетку. Нервные волокна меньшего размера образуют наружный кутикулярный слой, сосредоточенный вокруг сумки терминальных фолликулов и луковицы веллусных фолликулов. Вокруг волосяного фолликула расположено несколько типов нервных окончаний, включая свободные нервные окончания, ланцетовидные нервные окончания, клетки Меркеля и корпускулы пило-Руффини. Каждое нервное окончание воспринимает различные воздействия и стимулы. Свободные нервные окончания передают боль, ланцетовидные нервные окончания воспринимают ускорение, клетки Меркеля — давление, а структуры пило-Руффини — натяжение.
Перифолликулярные нервы содержат нейромедиаторы и нейропептиды, такие, как субстанция Р или пептид, связанный с кальцитониновым геном, которые воздействуют на фолликулярные кератиноциты, изменяя цикл волосяного фолликула Со своей стороны кератиноциты волосяных фолликулов вырабатывают нейротропные факторы, влияющие на перифолликулярные нервы и стимулируют их ремоделирование в зависимости от волосяного цикла. Клетки Меркеля, которые относят к нейроэндокринным клеткам, также синтезируют нейротропные факторы, цитокины и другие регуляторные молекулы. Поскольку клетки Меркеля сосредоточены в области волосяной сумки, некоторые авторы считают, что секретируемые ими факторы могут влиять на цикл волосяного фолликула.
е) Перифолликулярное влагалище. Перифолликулярное влагалище окружает эпителиальные компоненты волосяного фолликула и состоит из внутренней базальной мембраны, которая называется «гиалиновой мембраной», и наружного соединительнотканного влагалища. Базальная мембрана фолликула является продолжением межфолликулярной базальной мембраны. Она наиболее выражена вокруг наружного корневого влагалища у луковиц анагеновых волос. Во время катагена базальная мембрана утолщается и затем распадается.
Базальную мембрану окружает соединительнотканное влагалище, изначально состоящее из коллагена типа III. Вокруг верхнего фолликула имеется тонкое соединительнотканное влагалище, служащее продолжением окружающего сосочкового слоя дермы и расположено вертикально. Вокруг нижнего фолликула соединительнотканное влагалище выражено сильнее и имеет внутренний слой коллагеновых волокон, охватывающих фолликул, окруженный слоем продольно ориентированных коллагеновых волокон.
При трансплантации под кожу, эта перифолликулярная соединительная ткань обладает удивительной способностью образовывать новый дермальный сосочек и инициировать формирование нового волосяного фолликула. Даже если соединительнотканное влагалище трансплантируют другому человеку, эти фолликулы выживают без признаков иммунологического отторжения.
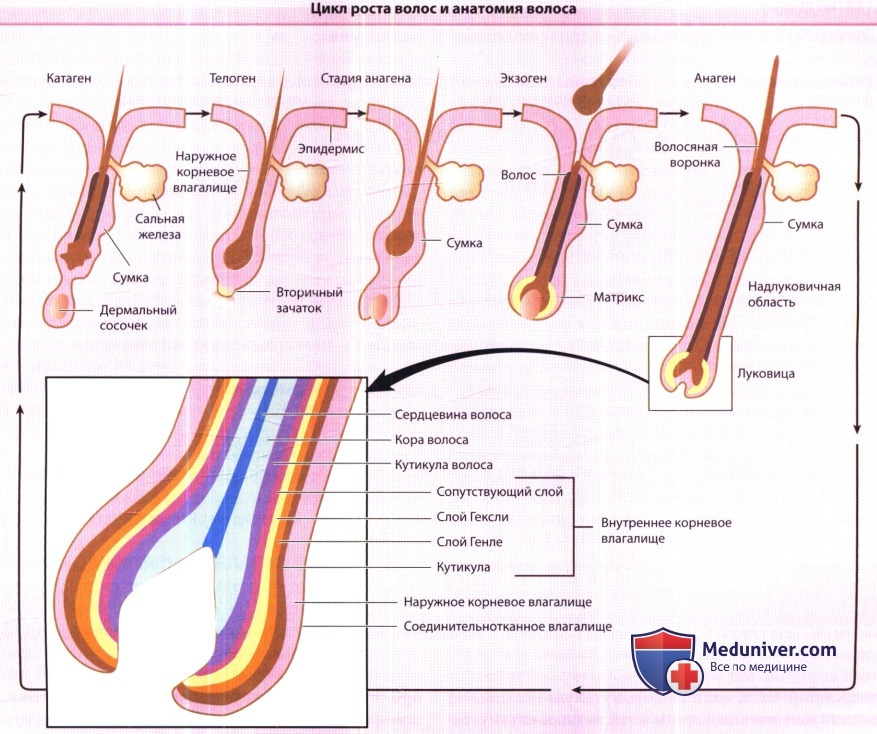
Цикл волосяного фолликула состоит из стадий покоя (телоген), роста волос (анаген), регрессии фолликула (катаген) и потери волоса (экзоген).
Вся нижняя эпителиальная структура формируется во время анагена и регрессирует во время катагена.
Переходная часть фолликула включает клетки матрикса в луковице, которые генерируют семь различных клеточных линий: три в стержне волоса и четыре во внутреннем корневом влагалище.
В = bulge (сумка); Е = epidermis (эпидермис); DP = dermal papilla (дермальный сосочек); Н = hair shaft (стержень волоса);
М = matrix (матрикс); ORS = outer root sheat (наружное корневое влагалище); S = sebaceous gland (потовая железа); Sec Grm = secondary germ (вторичный зачаток).
- Рекомендуем далее ознакомиться со статьей "Стадии роста волоса. Цикл волосяного фолликула"
Оглавление темы "Анатомия и физиология волос.":