
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Стадии роста волоса. Цикл волосяного фолликула
Каждый отдельный волосяной фолликул постоянно проходит через три стадии: (1) роста (анаген), (2) инволюции (катаген) и (3) покоя (телоген). Длительность анагена определяет окончательную длину волос и изменяется в зависимости от участка тела; длительность катагена и телогена менее вариабельна относительно участка тела. Самая длительная фаза анагена у волос на волосистой части кожи головы, она составляет от 2 до 8 лет. Длительность фазы анагена у молодых мужчин на других участках тела короче: на ногах — 5-7 месяцев, руках— 1,5-3 месяца, ресницах— 1-6 месяцев и на пальцах— 1-3 месяца.
В отличие от большинства млекопитающих, в том числе мышей и новорожденных детей, волосы на голове у взрослого человека растут асинхронно.
Примерно 90-93% фолликулов на волосистой части кожи головы находятся в фазе анагена, а остальные преимущественно в стадии телогена. Применив эти показатели к количеству волос на голове человека, которое составляет от 100000 до 150000 волосков, можно определить, что в любой момент времени в фазе телогена находятся 10000 волосков. Однако поскольку ежедневно мы теряем от 50 до 100 волосков, можно предположить, что стадия телогена является гетерогенным состоянием. Фолликулы, теряющие волосяной стержень, находятся, следовательно, в фазе «экзогена», которая охватывает примерно 1% телогеновых волосяных фолликулов. Волосы на голове растут со скоростью от 0,37 до 0,44 мм в день или примерно 1 сантиметр в месяц.
а) Стволовые клетки волосяного фолликула. Поскольку нижний участок фолликула циклически регенерируется, считалось, что этим ростом управляют фолликулярные стволовые клетки. Ранее предполагалось, что стволовые клетки находятся исключительно во «вторичном зародыше», расположенном у основания телогенового волосяного фолликула. Считалось, что в фазе анагена вторичный зародыш движется вниз к волосяной луковице и поставляет новые клетки для производства волоса.
Предполагалось, что в конце фазы анагена вторичный зародыш движется вверх с дермальным сосочком, чтобы в фазе катагена прийти в состояние покоя у основания телогенового фолликула. Этот сценарий движения стволовых клеток в ходе фолликулярного цикла был поставлен под вопрос, когда в зоне, окружающей фолликул телогенового волоса, была идентифицирована популяция долгоживущих, предположительно стволовых клеток. Затем было показано, что вторичный зародыш является переходной структурой, которая образуется в конце катагена из клеток нижнего вздутия (волосяной сумки). Концепция о постоянном нахождении стволовых клеток фолликула в этой волосяной сумке сейчас подтверждена в ходе анализа клеточных линий, который показал, что из клеток сумки происходят все эпителиальные слои волосяного фолликула. С этим согласуется и тот факт, что абляция клеток сумки приводит к деструкции фолликула.
Эти данные свидетельствуют в пользу концепции о том, что потеря стволовых клеток в сумке волосяного фолликула приводит к перманентной или рубцующей алопеции.
Достигнут прогресс в идентификации разновидностей клеток в пределах волосяного фолликула, которые относятся к различным популяциям стволовых и прогениторных клеток. Выявленные при анализе наследования сцепленных генов маркеры, способствующие постоянному возобновлению цикла волосяного фолликула, включают цитокератин 15 и Lgr5. Несмотря на то, что Lgr5 часто объявляется исключительным маркером вторичных зародышевых клеток, он является также маркером клеток волосяной сумки. Lgr6, ген, родственный Lgr5, экспрессирован на участке над волосяной сумкой в верхней части истмуса. Клетки, маркируемые Lgr6, мигрируют в эпидермис во время гомеостаза и после повреждения. Вдобавок к указанным, несколько других маркеров демонстрируют гетерогенность эпителия волосяного фолликула.
Являются ли клетки сумки «первичными» стволовыми клетками в эпителии кожи? Генерируют ли они, например, эпидермис и сальные железы во время гомеостаза и после ранения? Для ответа на эти вопросы применялись анализ клеточных линий и трансгенные методики. Как продемонстрировано на рисунке ниже, клетки волосяной сумки обычно не перемещаются в эпидермис, но после эксцизии кожи на полную толщину потомство клеток сумки мигрирует в рану во время реэпителизации. Эти клетки составляют примерно 30% клеток в регенерировавшем эпидермисе. Роль клеток сумки в поддержании жизнедеятельности сальной железы еще не ясна и находится в процессе изучения.
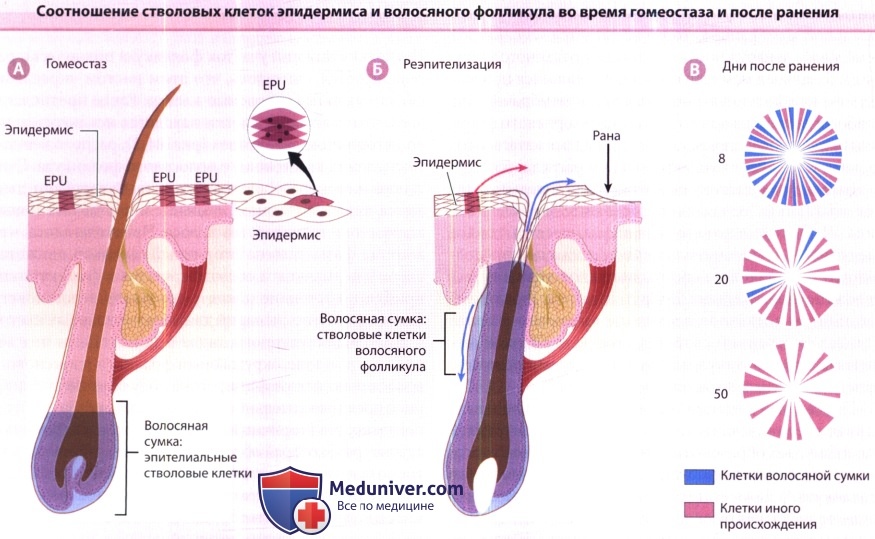
А. В обычных условиях обновление эпидермиса зависит от пролиферации клеток в эпидермальных пролиферативных единицах (EPU),
которые представляют собой клональные популяции клеток, организованные в колонки в форме шестигранника, которые производят одну наружную чешуйку.
Эпителиальные стволовые клетки из сумки волосяного фолликула не участвуют в обновлении эпидермиса.
Б. После ранения кожи на полную толщину, клетки волосяной сумки поступают в эпидермис для немедленного закрытия раны (синяя стрелка вверх).
Клетки волосяной сумки необходимы также для цикла волосяного фолликула (синяя стрелка вниз).
В. С течением времени количество клеток, происходящих из волосяной сумки, уменьшается,
и в ране после реэпителизации преобладают клетки иного происхождения (межфолликулярного эпидермиса и волосяной воронки).
б) Анаген. Процесс образования нижнего фолликула и волоса в начале фазы анагена аналогичен эмбриональному фолликулогенезу. Анаген можно разделить на семь этапов:
(1) этап I — рост дермального сосочка и начало митотической активности в расположенных сверху клетках эпителия, напоминающих зародышевые клетки;
(2) этап II — окружение матриксными клетками луковицы дермального сосочка и начало дифференцировки, формирующаяся луковица начинает движение вниз по фиброзному «серпантину»;
(3) этап III — начало дифференцировки матриксных клеток луковицы во все компоненты фолликула;
(4) этап — IV—реактивация матриксных меланоцитов;
(5) этап V—появление волосяного стержня и смещение телогенового волоса;
(6) этап VI — вход нового волосяного стержня на поверхность кожи и
(7) этап VII — стабильный рост.
В ходе пролиферации и миграции кератиноцитов в дерму для повторного образования нового нижнего фолликула на основном участке роста повляются такие ферменты, как коллагеназы и протеазы, а активность факторов роста и их рецепторов повышается, как и при ранении эпителия. Активируются также механизмы дифференцировки кератиноцитов, наблюдающиеся в эпидермисе во время заживления раны, в частности, экспрессия кератина 6. У мышей с отсутствием Stat3, регулятора миграции клеток в эпителии кожи, наблюдаются дефекты при заживлении ран и неспособность волосяных фолликулов войти в фазу анагена, что еще раз показывает сходство между заживлением раны и ранними процессами фазы анагена. Интересно отметить, что дермальный сосочек выживает в эпицентре этой деградирующей среды и движется вниз. Ремодулируются также сосудистая и иннервационная сети кожи.
Меланоциты пролифелируют и заселяют новую волосяную луковицу и наконец, эндотелиальная пролиферация и ангиогенез в дермальном сосочке обозначают тот момент, когда нижний фолликул полностью восстанавливается и активно производит новый волосяной стержень.
в) Катаген. Начало катагена обозначено прекращением митотической активности матриксных клеток и четко скоординированным апоптозом на циклическом участке волосяного фолликула. Выработка пигмента меланоцитами прекращается до остановки пролиферации матриксных клеток, что приводит к отсутствию пигментации на проксимальном конце клубкового телогенового волоса. Меланин часто обнаруживается в соседней дерме и сосочке, там, где он окружен макрофагами. Перифолликулярное влагалище распадается и, гиалиновая мембрана утолщается. Нижний фолликул оттягивается вверх вместе с дермальным сосочком. Перифолликулярное влагалище образует фиброзный «серпантин», состоящий из фибробластов, мелких кровеносных сосудов и коллагена. В итоге дермальный сосочек располагается непосредственно под волосяной сумкой в нижней части перешейка.
В фазе катагена самые крупные фолликулы, например, на волосистой части кожи головы, уменьшают свою длину до 2-5 миллиметровых структур, самая глубокая часть которых, луковица, достигает подкожно-жировой клетчатки, до укороченных фолликулов, размеры которых в телогене составляют от 0,25 до 0,5 мм. По мере утолщения базальной мембраны вокруг нижнего фолликула дермальный сосочек, защищенный от окружающего апоптоза и деструкции (возможно потому, что экспрессирует Bcl-2, анти-апоптотический фактор), конденсируется и начинает двигаться вверх, чтобы в фазе телогена перейти в состояние покоя на участке под волосяной сумкой.
Для непрерывности фолликулярного цикла необходима миграция дермального сосочка от подкожно-жировой клетчатки в дерму во время фазы катагена. Иллюстрацией этому может служить синдром атрихии с образованием папул. У таких пациентов отмечаются мутации либо в их безволосом гене, либо в гене рецептора витамина D при рахите. У мышей с подобными мутациями наблюдается безволосый фенотип. На примере таких мышей стало понятно, что фолликулогенез в данном случае протекает нормально, однако когда фолликулы входят в фазу катагена в первый раз, нижняя часть фолликула не подвергается должной инволюции и сокращению, и дермальный сосочек остается изолированным в подкожно-жировой клетчатке. Хотя клетки волосяной сумки все еще присутствуют, новых анагеновых фолликулов так и не образуется, предположительно потому, что стволовые клетки не могут вступить во взаимодействие с дермальным сосочком.
Изучение мутантных мышей привело к нескольким ключевым открытиям, которые углубили наше понимание молекулярных явлений в начале фазы катагена. В частности, Herbert и соавт. обнаружили, что у мышей с отсутствием гена Fgf5 волосы на 50% длиннее, чем у обычных диких мышей, и что мутации в этом гене отвечают за фенотип ангоры, который был описан более 30 лет назад. Хотя эти результаты оказались довольно неожиданными, тщательное изучение экспрессии Fgf5 на протяжении нормального цикла роста волос показало, что экспрессия гена активируется в наружном корневом влагалище и клетках волосяного матрикса непосредственно перед началом катагена, следовательно, Fgf5 способен запускать начало фазы катагена. Интересно, что фолликул все-таки в итоге переходил в фазу катагена даже в отсутствие FGF-5, что указывает на избыточность в механизме FGF-5 или на особую, присущую матриксным клеткам пролиферативную способность.
Дальнейшие исследования также показали, что другие члены семейства FGF и их рецепторы экспрессированы во время анагена и, вероятно, также играют некоторую роль в цикле развития волосяного фолликула. Фенотип волосяного покрова у мыши с дефицитом FGF-5 существенно изменяется при эктопической экспрессии анти-апоптотического гена bcl-xLX в наружном корневом влагалище, что указывает на возможную роль наружного корневого влагалища в контроле над циклом роста волос.
Хотя многие годы было известно, что экзогенное назначение EGF овцам приводило к индукции катагена, но только в результате последних исследований трансгенных и нокаутных мышей стала понятной важность системы EGFR в регулировке цикла роста волос.
Например, у мышей, нокаутных по трансформирующему фактору роста волос-а (TGF-а), основному лиганду EGFR, отмечается аномальное развитие волосяного фолликула и проявляется волнистый фенотип роста волос, если EGFR функционально деактивируется в базальном слое эпидермиса и волосяном фолликуле с применением доминантной негативной трансгенной стратегии. Возникающий в результате волос не только вьется, но и длиннее, чем нормальный. Переход волосяных фолликулов из анагена в катаган у этих мышей запаздывает. Волосяные фолликулы в коже мыши, у которой EGFR полностью отсутствует, также не переходят из анагена в телоген. Таким образом, для нормального развития волосяного фолликула и его цикла требуется наличие EGFR и его лиганда.
Учитывая сложный состав семейства EGFR, к которому относятся четыре рецептора (ErbB1—4) и как минимум шесть лигандов, необходимы дальнейшие исследования для выяснения роли индивидуальных членов этого семейства в цикле волосяного фолликула.
Кроме FGF-5 и EGF, преждевременный катаген вызывают нейротропины и TGF-βl. Нейротропин-3 и нейротропный фактор головного мозга у трансгенных мышей демонстрируют преждевременное развитие фазы катагена, а избыточная экспрессия нейротропного фактора головного мозга приводит к сокращению длины волос на 15%, вероятно, вследствие стимуляции проапоптотической сигнализации через рецептор нейротропина p75kd. TGF-1 индуцирует преждевременный катаген в изолированных волосяных фолликулах человека и в коже мышей in vivo, а у мышей, нокаутных по TGF-β1, начало фазы катагена замедленно.
г) Телоген и экзоген. После завершения инволюции катагена и формирования клубкового волоса волосяной фолликул готовит волос для отторжения с волосистой части кожи головы. Каждый день отторгается примерно 1% волосяных фолликулов. Millner и соавт. предложили выделить утрату волос в отдельную фазу, которую назвали экзогеном. У млекопитающих, которые линяют на сезонной основе фаза экзогена строго контролируется и имеет жесткие временные рамки. Тот факт, что экзоген является активной стадией, подтверждается исследованиями Хедингтона (Headington), который описал тип выпадения волос, обозначенным термином «немедленное телогеновие высвобождение».
Такой тип потери волос может наблюдаться вскоре после приема лекарственных препаратов, таких как миноксидил, или же в ответ на быструю смену света и темноты. Процесс состоит в усиленном выпадении клубковых волос в течение недель после провоцирующего события (слишком быстро, чтобы быть вызванными фолликулами, преждевременно вошедшими в фазу телогена из анагена), что указывает на возможность активной утраты клубковых волос, которые в норме сохраняются в фолликуле. Факт гетерогенности телогена находит подтверждение в работе Guarrera и Reborra, которые проследили поведение индивидуальных волос in situ, используя макрофотографии, в течение более двух лет и показали, что между утратой и повторным ростом волос может пройти несколько месяцев. Такой «период запаздывания» в норме не наблюдается или же бывает очень коротким, но у пациентов с андрогенетической алопецией часто продолжается несколько месяцев.
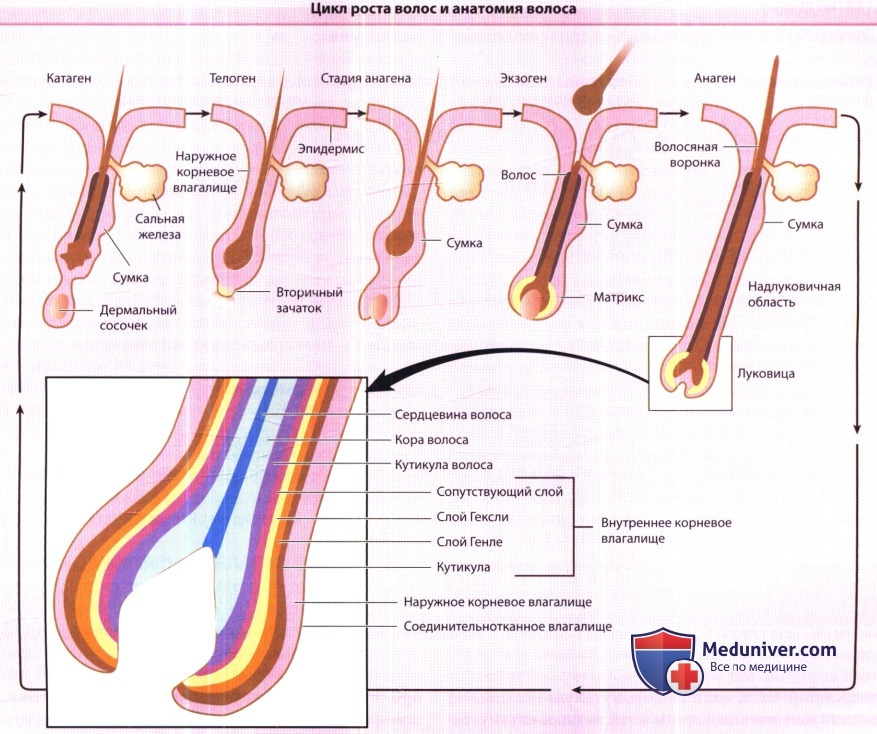
Цикл волосяного фолликула состоит из стадий покоя (телоген), роста волос (анаген), регрессии фолликула (катаген) и потери волоса (экзоген).
Вся нижняя эпителиальная структура формируется во время анагена и регрессирует во время катагена.
Переходная часть фолликула включает клетки матрикса в луковице, которые генерируют семь различных клеточных линий: три в стержне волоса и четыре во внутреннем корневом влагалище.
В = bulge (сумка); Е = epidermis (эпидермис); DP = dermal papilla (дермальный сосочек); Н = hair shaft (стержень волоса);
М = matrix (матрикс); ORS = outer root sheat (наружное корневое влагалище); S = sebaceous gland (потовая железа); Sec Grm = secondary germ (вторичный зачаток).
- Рекомендуем далее ознакомиться со статьей "Механизм пигментации волос. Регуляция цвета волос"
Оглавление темы "Анатомия и физиология волос.":