
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Эмбриология волос и ее регуляция
- Главным предназначением волос для людей является их влияние на социальные взаимоотношения.
- Развитие волосяного фолликула зависит от взаимодействий между эпителиальными и мезенхимными клетками. Важные для этого взаимодействия гены постепенно идентифицируют.
- В цикле возобновления волосяного фолликула определенная роль принадлежит генам, определяющим развитие фолликулов.
- В сумке волосяного фолликула содержатся стволовые клетки, необходимые для цикла непрерывной регенерации фолликула.
- Пигментация волос зависит от меланоцитарных стволовых клеток и дифференцированных клеток в фолликуле. Идентифицировано несколько генов, определяющих поведение меланоцитов и пигментацию волос.
Волосяной покров имеется только у млекопитающих. В ходе эволюции ему отводилась роль изоляции и защиты от внешней среды. Однако у современного человека основное предназначение волос сводится к их роли в социальных взаимоотношениях. Потеря волос (алопеция), как и избыточный рост в нежелательных участках (гирсутизм и гипертрихоз) приводят к значительному психологическому и эмоциональному стрессу, которым оправдываются дорогостоящие усилия, направленные на то, чтобы обратить эти состояния вспять.
Продолжает углубляться фундаментальное понимание процесса роста волос и контролирующих его факторов, что приводит к появлению новых методов лечения алопеции. Этот прогресс был вызван интересом исследователей в области биологии развития и смежных отраслей к волосяному фолликулу, как модели для широкого спектра биологических процессов.
По мере того как каждый волосяной фолликул циклически регенерирует, он повторяет свое первоначальное развитие. Многие факторы роста и рецепторы, играющие важную роль во время развития волосяного фолликула, регулируют также и циклическую регенерацию фолликула. В состав волосяного фолликула входят стволовые клетки кератиноцитов и меланоцитов, нервы и сосудистая сеть, которые важны как для здоровой, так и для больной кожи. Для применения появившейся информации и надлежащего осмотра пациента с потерей или избыточным ростом волос большое значение имеет понимание анатомии и развития волосяного фолликула.
а) Эмбриология роста волос. Морфологически развитие волосяного фолликула подразделяется на восемь последовательный стадий, некоторые из них представлены на рисунке ниже. Для каждой стадии характерна уникальная картина экспрессии факторов роста, их рецепторов и антагонистов, молекул адгезии и компонентов внутриклеточной сигнализации. Прогресс в понимании молекулярных механизмов развития волосяного фолликула был достигнут благодаря открытию того, что гены млекопитающих, гомологичные генам, ответственным за нормальное развитие плодовой мухи дрозофилы, влияют также и на развитие волосяного фолликула.
Решающее значение для развития волосяного фолликула и позвоночных в целом имеют такие гены, как декапентоплегический комплекс [Dpp/bone morphognenetic protein (BMP)], ген engrail (en), семейство гомеобоксных генов (hox), а также гены hh/ptc (hedgehog-patched), notch и wg/wnt/catenin (wingless/armadillo). Все эти гены были впервые открыты у дрозофилы, и большинство данных им названий носит описательный характер, касающийся специфической внешности (фенотипа) дрозофил, несущих мутации в данных генах.
Формирование фолликулов in utero начинается на голове, а затем продолжается на остальных участках тела. Вначале образуется зародышевый пушок — мягкие, тонкие, лишенные пигментации первичные волосы плода. Первичный пушок обычно отпадает между 32 и 36 неделями развития плода, хотя примерно у одной трети новорожденных он сохраняется еще несколько месяцев после рождения.
Паттернирующие гены, которые называют «гомеобоксными генами», имеют точную геномную организацию, так что в ходе развития они экспрессированы в строго определенных как временных последовательностях, так и пространственных расположениях. Эти гены, вероятно, ответственны за неслучайное и симметричное распределение волосяных фолликулов по телу. у взрослых мышей экспрессия гомеобоксных генов вновь проявляется в волосяных фолликулах и служит для поддержания нормального воспроизводства волосяных стержней.
Ген «engrailed», принадлежащий к типу гомеобоксных генов, отвечает за дорсально-вентральную паттернизацию, поэтому у мышей с отсутствием этого гена волосяные фолликулы развиваются на подушечках лапок.
Хотя базовая анатомия волосяных фолликулов и волос одинакова, их рост, размер, форма, пигментация и другие характеристики существенно отличаются в зависимости от локализации на теле и варьируют у различных индивидуумов. Многие из перечисленных характеристик формируются в ходе развития, а затем значительно изменяются вследствие гормональных влияний. Мы начинаем понимать, какие гены контролируют длину волос, их курчавость и распределение, благодаря отличным генетическим исследованиям на собаках.
Эти исследования выявили, что на длину, курчавость и распределение волос влияют соответственно фактор роста фибробластов-5 (FGF-5), кератин 71 и R-спондин 2. У людей, более толстые стрежни волос у азиатов связаны с повышенной активностью рецептора эктодисплазина (EDAR), о рецепторе эктодисплазина (EDA) речь пойдет ниже.
Размер многих типов фолликулов существенно изменяется несколько раз в течение жизни. Например, волосы зародышевого пуха (лануго), длиной в несколько сантиметров, преобразуются в фолликулы веллусных волос, которые производят небольшие волоски, лишь слегка выступающие над поверхностью кожи. В более поздний период веллусные фолликулы в области роста бороды у мужчин укрупняются до терминальных фолликулов, которые производят толстые длинные волосы. Фолликулы терминальных волос на волосистой части кожи головы у генетически предрасположенных индивидуумов резко уменьшаются и производят слабые микроскопические волоски.
б) Эпителиальная плакода или первичный зародыш волоса. У эмбриона человека волосяные фолликулы развиваются из небольших скоплений клеток, которые называются эпителиальными плакодами и соответствуют первой стадии развития волосяного фолликула. Они впервые появляются на 10 неделе беременности. Затем эпителиальная плакода расширяется и образует «первичный зародыш волоса», из которого развивается вся эпителиальная часть волосяного фолликула.
Клетки волосяной плакоды и зародыша экспрессируют плацентарный кадгерин и принимают вертикальную ориентацию, теряя свои десмосомы, полудесмосомы и эпителиальный кадгерин, что уменьшает их адгезию с соседними клетками. Дермальные клетки под волосяной плакодой образуют кластер (или конденсат), который затем развивается в дермальный сосочек.
Формирование волосяного фолликула зависит от ряда взаимодействий между мезенхимой и эпителием. Первоначальный сигнал возникает в мезенхиме (первичной дерме) и стимулирует расположенный над ней эпителий к образованию придатка, на что указывает появление равномерно расположенных плакод. Второй сигнал идет от эпителиальной плакоды и приводит к скоплению клеток в подлежащей мезенхиме, из которых затем формируется дермальный сосочек. И наконец, сигнал от этого первичного дермального сосочка инициирует пролиферацию и дифференцировку клеток плакоды, что в итоге приводит к образованию зрелого фолликула.
Эти взаимные сигналы проходят через промежуточную базальную мембрану, морфология и химический состав которой изменяется, что влияет на способность мембраны секвестировать факторы роста и связывающие белки и, возможно, модулирует, таким образом, эпителиально-мезензхимные взаимодействия.
Идентифицированы несколько регуляторных молекул, необходимых для образования волосяного фолликула, но еще предстоит определить, как они взаимодействуют, чтобы генерировать волосяные фолликулы в гомогенном эпителии. В одной из моделей расположение и размер плакод регулируются дермальным сигналом, характер которого варьирует в различных участках тела. Дермальный сигнал поступает одинаково во все участки тела и запускает активацию промоутеров и репрессоров фолликулов, которые затем конкурируют друг с другом, что приводит к появлению соответствующего набора фолликулов.
Региональные различия в размере и расположении фолликулов объясняются разницей в уровнях активации промоутеров и репрессоров. В соответствии с этой моделью несколько положительных и отрицательных регуляторов судьбы волосяного фолликула вначале равномерно экспрессированы в эпидермисе, а затем локализуются в плакоды.
Одним из самых ранних молекулярных механизмов, положительно регулирующих инициацию волосяного фолликула, является путь WNT/β-катенин. β-катенин служит медиатором нижних звеньев WNT-сигнализации. WNT-белки связываются с рецепторами на клеточной мембране и через серию сигналов тормозят деградацию цитоплазматического β-катенина. β-катенин транслоцируется в ядро, образуя комплекс с семейством факторов транскрипции LEF/TCF, что приводит к экспрессии генов, регулирующих нижележащие (относительно контрольных точек клеточного цикла) звенья сигнальных каскадов.
Активация данного β-катенинового пути необходима для возникновения эпителиальной компетенции — состояния, при котором эпителиальная ткань обладает потенциалом формирования волосяного фолликула. Обычно во взрослом эпителии β-катениновый путь находится в неактивном состоянии, однако при искусственной активации β-катенина в базальных клетках эпидермиса взрослых трансгенных мышей волосяные фолликулы развиваются de novo. Этот примечательный результат, возможно, будет использоваться в терапевтических разработках, хотя постоянная активация данного пути в волосяном фолликуле приводит также к появлению пиломатриком и трихофолликулом, двух типов сравнительно редких опухолей кожи.
Эктодиплазин (EDA) — молекула, родственная фактору некроза опухоли,— и его рецептор (EDAR) также являются частью еще одного главного пути, который стимулирует раннее развитие фолликулов как у мыши, так и у человека. Мутации в гене EDA вызывают Х-сцепленную ангидротическую эктодермальную дисплазию, синдром, который ассоциируется с уменьшением количества волосяных фолликулов, дефектами зубов и потовых желез. Мутации в гене EDAR отмечаются при аутосомно-рецессивных и доминантных гипогидротических эктодермальных дисплазиях, вызывающих фенотипы, идентичные фенотипам вследствие мутаций EDA.
Мышиный ген Edar экспрессирован в эпителии до образования плакоды повсеместно, а затем его экспрессия ограничена теми плакодами, в которых ген Eda экспрессирован повсеместно, даже после формирования плакоды. У мышей с мутациями этих генов наблюдается тот же фенотип, что и у людей с подобными мутациями, а избыточная экспрессия Eda в эпителии мышей приводит к образованию «сливных» фолликулов вследствие утраты соответствующих интервалов между соседними волосяными плакодами. У лиц с большей активностью генов EDAR волосы толще.
В отличие от EDA и EDAR, которые стимулируют развитие волосяного фолликула, члены семейства ВМР тормозят его формирование. Bmp2 диффузно экспрессирован в эктодерме, а затем локализуется в раннюю плакоду и подлежащую мезенхиму, в то время как Bmp4 экспрессирован в раннем дермальном конденсате. ВМР-сигнализация тормозит образование плакоды, в то время как нейтрализация активности BMP антагонистом этого белка ноггином содействует ее развитию по крайней мере частично, через положительную регуляцию экспрессии фактора, связывающего лимфоидный энхансер 1 (Lef-1). При отсутствии ноггина волосяных фолликулов у мышей меньше нормы и развитие фолликулов замедленное.
Сигнальный путь «Notch» также влияет на картину развития фолликулов. Notch-лиганд-Δ1 в норме экспрессирован в мезенхиме, расположенной под плакодой и, будучи ошибочно экспрессированным в небольшом участке эпителия, стимулирует и ускоряет развитие плакоды, подавляя формирование плакоды в окружающих клетках.
Еще один секретируемый белок, присутствующий в фолликулярной плакоде и играющий важную роль в эпителиально-мезенхимной сигнализации — это Shh (sonic hedgehog). Кожа мышей при отсутствии Shh имеет очень слабые волосяные фолликулы с плохо развитыми дермальными сосочками.
Ptc1 (patched1), рецептор Shh, экспрессирован в зародышевых клетках и подлежащем дермальном сосочке, следовательно, Shh может обладать как аутокриновыми, так и паракриновыми индуктивными свойствами, необходимыми для формирования волосяного зародыша и дермального сосочка. Patched-ген является дефицитным при синдроме базальноклеточного невуса.
в) Волосяная почка. На следующей стадии формируется волосяная почка (стадия 2 развития волосяного фолликула), при этом волосяной зародыш удлиняется, образуя тяж эпителиальных клеток. Мезенхимные клетки с обеих сторон почки развиваются в фиброзное влагалище волосяного фолликула, а клетки у вершины почки образуют дермальный сосочек. Самый глубокий участок фолликулярной почки образует луковицеобразную структуру, окружающую подлежащие мезенхимные клетки, которые станут дермальным сосочком. Эти эпителиальные клетки образуют матрикс волосяного фолликула, из которого разовьются волосяной стержень и внутреннее корневое влагалище. На наружном корневом влагалище образуются два вздутия со стороны волосяного фолликула под тупым углом к поверхности кожи.
Поверхностное вздутие разовьется в сальную железу. Более глубокое вздутие — место локализации эпителиальных стволовых клеток, которые генерируют новый фолликул во время цикла развития волосяного фолликула. Мышца, поднимающая волос, обычно прикрепляется к зоне вздутия, и сокращение мышцы усиливает вертикальную ориентацию волосяного стержня, вызывая появление «гусиной кожи». В подмышечных впадинах, аногенитальной области, ареолах, околопупочной области, на веках (специализированные моллевые железы) и наружных слуховых проходах развивается третье вздутие выше почки сальной железы, из которого развивается апокриновая железа.
По мере формирования фолликулярной луковицы на стадии волосяной почки развивается как минимум восемь различных клеточных слоев, составляющих все компоненты зрелого волосяного фолликула. Понимание того, какие гены определяют специфические клеточные линии в пределах фолликула, является важным вопросом. GATA-связывающий белок-3 важен для дифференцировки внутреннего корневого влагалища. Notch1, мембранный белок, участвующий в определении судьбы клетки в процессе межклеточных взаимодействий и внутриклеточной передачи сигнала, и его лиганды Serratel и Serrate2 экспрессированы в матричных клетках, которые образуют внутреннее корневое влагалище и волосяной стержень. Notch 1, вероятно, контролирует фенотип кератиноцитов, когда они покидают луковичный матрикс и дифференцируются в специфические клеточные типы.
г) Зрелый волосяной фолликул. Центральный люмен, где появляется волосяной стержень, образуется вследствие некроза и ороговения эпителиальных клеток в волосяной воронке. В производстве волосяного стержня участвуют несколько сигнальных путей, которые контролируют процессы его дифференцировки. Важную роль в формировании волосяного стержня играет сигнальный путь Wnt/β-катенин/Lef-1, а эктопическая экспрессия Wnt3 в наружном корневом влагалище волосяного фолликула приводит к ломкости волосяного стержня. Гены кератина волосяного стержня содержат участки связывания Lef-1, который транслоцируется в ядро после активации пути WNT-β-катенин. WNT-сигнализация, возможно, регулирует экспрессию генов кератина волосяного стержня, поскольку почти все эти гены содержат связывающие Lef-1 участки в своих промоутерных регионах.
BMP-сигнализация также важна для соответствующей дифференцировки внутреннего корневого влагалища и волосяного стержня, поскольку делеция BMP-рецептора типа 1А в кератиноцитах приводит к значительным изменениям внутреннего корневого влагалища и процесса формирования стержня волоса. Дифференцировку волосяного стержня предположительно контролируют и некоторые другие факторы транскрипции, включая гомеобоксный белок НОХС13 и ген WHN, в котором происходят мутации у голых мышей и редко у людей с дефектами волос, ногтей и иммунитета.
Процесс образования волосяного фолликула повторяется волнообразно несколько раз, при этом наряду с первоначальным фолликулом образуются вторичные фолликулы. Фолликулы изначально собраны в группы по три и ориентированы наклонно под одинаковым углом к своим соседям.
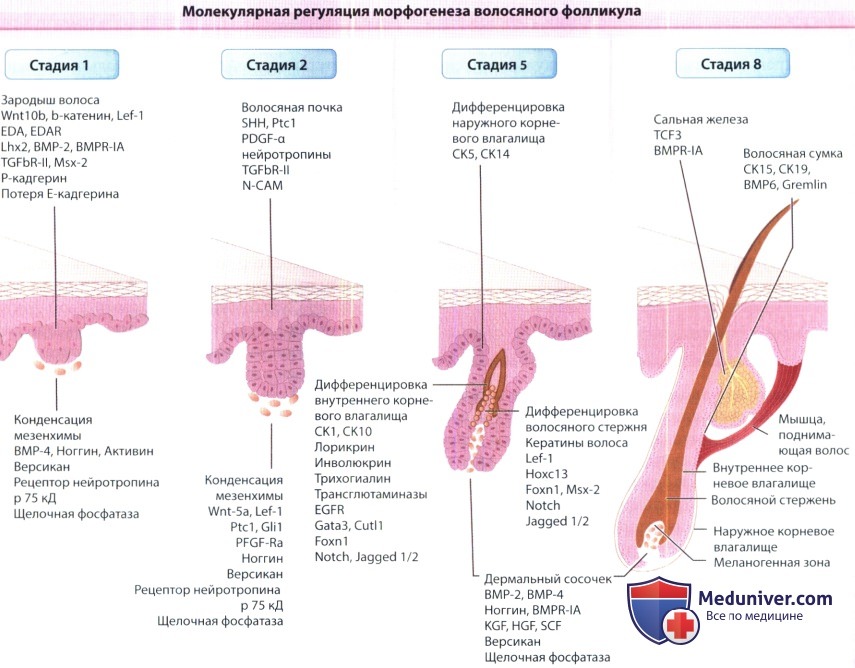
Схема показывает экспрессию различных факторов роста, их рецепторов, молекул адгезии и клеточного матрикса,
регуляторов транскрипции в фолликулярном эпителии и мезенхиме на различных стадиях развития волосяного фолликула.
BMP = морфогенный костный белок; BMPR-IA = рецептор морфогенного костного белка типа IA; СК = кератин;
Cutl1 = cut-like1; E-cadherin = эпителиальный кадгерин; ЕОА=эктодисплазин;
EADAR = рецептор эктодисплазина; EGFR = рецептор эпидермального фактора роста; Foxn1= forkheadbox N1;
Gata3 = GATA-связывающий белок 3; Gli1 =глиома-ассоциированный онкогенный гомолог 1; HGF = фактор роста гепатоцитов;
Hoxc13 = гомеобокс C13;KFG = фактор роста кератиноцитов; Lef-1 = фактор, связывающий лимфоидный энхансер; Lhx2 = ПМ-гомеобокс 2;
N-CAM = молекула адгезии нейральных клеток; P-cadherin = Р-кадгерин; PDGF-α = тромбоцитарный фактор роста α-полипептид;
PDGF-Rα = рецептор тромбоцитарного фактора роста α; Ptc1 = patched 1; SCF = фактор стволовых клеток;
Shh = sonic hedgehog; TCF3 = фактор транскрипции 3; TGF-βR-II = рецептор-2 трансформирующего фактора роста.
- Рекомендуем далее ознакомиться со статьей "Типы волос и их характеристика"
Оглавление темы "Анатомия и физиология волос.":