
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Тканевые базофилы. Кератинизация - образование рогового слоя.
В литературе имеются сведения о проникновении тканевых базофилов в интактный эпидермис. Так, К. Ш. Баймухамбетов и соавт. (1991) обнаружили миграцию этих клеток из дермы в эпидермис у тонкорунных овец. В эпидермисе тканевые базофилы подвергались дегрануляции, при этом со временем гранулы, вышедшие из клеток, постепенно светлели, вакуолизировались и исчезали. Есть сведения о наличии тканевых базофилов и в интактном эпидермисе человека. Данные наблюдения показывают, что сфера функциональных отправлений тканевых базофилов не ограничивается только соединительной тканью. Ранее описано проникновение тучных клеток в мерцательный эпителий дыхательных путей и в многослойный плоский эпителий шейки матки.
В работе О. Д. Мядельца (1993) установлен факт проникновения тканевых базофилов в многослойный эпителий новообразующихся волосяных фолликулов при органотипической регенерации кожи. При мастоцитозе тканевые клетки проникают в эпидермис в большом количестве.
Морфология процесса кератинизации была описана достаточно хорошо еще на уровне светового микроскопа. В ходе этого процесса кератиноциты увеличиваются в размерах (до 30 раз), уплощаются, в них постепенно конденсируется ядро, в цитоплазме накапливается кератогиалин. Затем ядро подвергается разрушению, а цитоплазма гомогенизируется. В клетках накапливается кератин. В конце концов клетки превращаются в роговые чешуйки. Электронный микроскоп позволил установить более тонкие процессы, происходящие при кератинизации. После миграции базального кератиноцита в шиповатый слой увеличивается число десмосом, связывающих его с другими клетками, а также численность тонофиламентов.
В верхних рядах шиповатого слоя в кератиноцитах появляются пластинчатые гранулы Одланда. Гранулы Одланда занимают периферическое положение и выделяют путем экзоцитоза свое содержимое в межклеточное пространство, где оно приобретает пластинчатое строение (межклеточный цемент). В цитоплазме зернистых кератиноцитов выявляются тонофибриллярно-кератогиалиновые комплексы. Цитолемма клеток заметно утолщается. В ядрах клеток часто видны глубокие инвагинации ядерной оболочки. Хроматин распределяется неравномерно, образуя главным образом скопления около ядерной мембраны. В нуклеоплазме нарастают явления разрежения или просветления. В конечном итоге ядра клеток разрушаются и исчезают.
Постепенно редуцируются органеллы. Матрикс митохондрий разрежается, в них уменьшается число крист. В последующем митохондрии разрушаются и исчезают. Исчезают эндоплазматическая сеть и свободные рибосомы. Кератиноциты превращаются в роговые чешуйки, склеенные цементирующим материалом и десмосомами. Десмосомы при этом видоизменяются: в них теряется межклеточный компактный слой с последующим слиянием промежуточных слоев. Роговые чешуйки в роговом слое различаются по распределению кератиновых фибрилл, которое постепенно изменяется по мере созревания роговых чешуек от менее к более плотному.
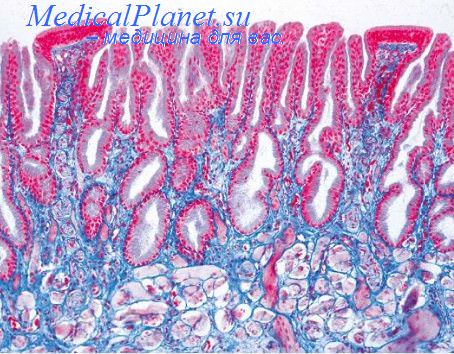
Биохимические и иммуноцитохимические исследования лишь в последнее время позволили понять многие тонкие механизмы кератинизации. Синтез кератина в форме прекератина начинается в базальных кератиноцитах. Прекератин имеет более низкий молекулярный вес, чем зрелый кератин (40—60 000 дальтон при молекулярной массе зрелого кератиноцита более 60 000 дальтон) и характеризуется отсутствием внутри- и межцепочечных дисульфидных связей, придающих молекулам кератина прочность и нерастворимость. В более высоких слоях эпидермиса (шиповатый, зернистый) биосинтез прекератина продолжается и даже нарастает, причем молекулярная масса синтезируемого прекератина увеличивается. Кератин теряет свойство растворимости в воде и приобретает прочность.
Для упаковки кератина необходим особый белок филагрин (ранее он назывался «белок, богатый гистидином». Этот белок вызывает агрегацию кератиновых филаментов. Биосинтез филагрина осуществляется в зернистом слое с участием специфической и-РНК. Филагрин обладает кислотными свойствами и отличается сильной степенью фосфорилирования. По мере синтеза он накапливается в виде кератогиалиновых гранул и существует до тех пор, пока плотно упакованный кератин не стабилизируется прочными дисульфидными связями. Как только это произойдет, филагрин в корнеоцитах рогового слоя распадается до свободных аминокислот.
Интересна их функция в корнеоците: свободные аминокислоты поддерживают в нем на высоком уровне осмотическое давление, что вызывает приток воды, которая сохраняется в роговых чешуйках даже при понижении влажности внешней среды. Аккумулированная вода придает чешуйкам упругость и механичесую прочность. Кроме того, аминокислота гистидин превращается в роговых чешуйках в урокановую кислоту, участвующую в защите кожи от ультрафиолетовых лучей. Все эти превращения филагрина и входящих в него аминокислот происходят при участии ферментов, что противоречит укоренившемуся представлению о роговой чешуйке как о мертвой клетке.
В клетках зернистого слоя синтезируется также белок кератолинин или инволюкрин. Этот белок выстилает внутреннюю поверхность цитолеммы зернистых кератиноцитов, утолщая ее. Утолщенная плазмолемма успешно противостоит гидролитическим процессам, которые интенсивно протекают в верхних рядах клеток зернистого слоя и приводят к разрушению кератиноцитов. В роговом слое молекулы кератолинина сшиваются глютамил — лизиновой связью при помощи ферментов трансглютаминаз и дисульфидными мостиками при помощи кальцийзависимой сульфгидрилоксидазы. Появление этих связей в белковых молекулах обусловливает прочность и нерастворимость их даже в очень сильных реагентах.
В результате корнеоцит приобретает прочную толстую так называемую поперечно-сшитую оболочку, характерную только для ороговевающего эпителия. У ряда наземных животных не обнаруживаются кератогиалиновые гранулы, а у некоторых водных млекопитающих, кроме того, не образуется полноценная поперечно сшитая оболочка. Аналогичная ситуация наблюдается при ряде заболеваний кожи.
В процессе кератинизации важная роль принадлежит кератиносомам. Эти видоизмененные лизосомы путем экзоцитоза выделяют свое содержимое в межклеточные пространства, где оно превращается в богатый липидами межклеточный цемент с пластинчатым строением. Если корнеоцит содержит лишь 3 % липидов, то межклеточный цемент — 80 % (остальные 20 % приходятся на белки различной молекулярной массы). Кератиносомы содержат по крайней мере 4 группы веществ: гликолипиды, гликопротеины, гидролитические ферменты, свободные стерины. По биохимическим данным в межклеточном матриксе обнаруживаются церамиды, холестерин и его эфиры, свободные жирные кислоты, триглицериды, сквалены, белки.
Основную массу составляют церамиды. Все эти вещества обеспечивают адгезию корнеоцитов, а также, за счет гидрофобное™ своих молекул, формируют барьерный слой эпидермиса.
Таким образом, процессы, происходящие при кератинизации, заключаются в следующем: образовании нерастворимого белка кератина; видоизменении десмосом с растворением межклеточного слоя; лизисе ядер кератиноцитов и их органелл; экзоцитозе гранул Одланда с образованием гидрофобного межклеточного цементирующего матрикса; образовании кератолинина, укреплении его глутамил-лизиновыми и дисульфидными связями с формированием нерастворимой поперечно-сшитой оболочки; гибели клетки и превращении ее в роговую чешуйку или корнеоцит.
- Читать далее "Эпидермальная пролиферативная единица. Гистология эпидермальной пролиферативной единицы."
Оглавление темы "Гистология и физиология эпидермиса.":1. Клетки Гринстейна. Меланоциты.
2. Клетки Меркеля. Функции и физиология клеток Меркеля.
3. Роль клеток Меркеля в патологии. Гистопатология клеток Меркеля.
4. Лимфоциты в коже. Функции и физиология лимфоцитов в коже.
5. Тканевые базофилы. Кератинизация - образование рогового слоя.
6. Эпидермальная пролиферативная единица. Гистология эпидермальной пролиферативной единицы.
7. Гомеостаз в эпидермисе. Обновление клеток в эпидермисе.
8. Генетический контроль дифференциации клеток эпидермиса. Гормональный контроль клеток в эпидермисе.
9. Аутокринный контроль клеток эпидермиса. Фактор роста эпидермиса.
10. Деление кератиноцитов. Пути миграции клеток эпидермиса - кератиноцитов.