
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Альтернативный путь системы комплемента. Фагоцитирующие клетки.
Активация комплемента по альтернативному пути происходит без участия АТ и задолго до их появления. Факторы альтернативного пути имеют буквенное обозначение: Р (пропердин), В и D (ферменты системы комплемента). Запуск альтернативного пути осуществляет активация компонента СЗ, взаимодействующего с факторами В и D. Затем через образование компонента С5 (но без участия С1, С2 и С4) альтернативный путь также завершается образованием на поверхности клеток-мишеней мембраноповреждающего комплекса. Альтернативный путь активируется такими микробными продуктами, как эндотоксины бактерий, вирусы (например, гриппа).
Фагоцитирующие клетки.
Фагоциты выполняют не только защитные (поглощают и разрушают чужеродные агенты), но и дренажные функции (удаляют погибшие и деградировавшие структуры организма). Фагоциты представлены клетками миелопоэтического ряда (полиморфноядерные лейкоциты) и макрофагально-моноцитарной системы (моноциты, тканевые макрофаги). Основные свойства фагоцитирующих клеток представлены в таблице.
Фагоцитоз — процесс поглощения и переваривания фагоцитами микроорганизмов, других клеток, фрагментов некротизированной ткани и чужеродных частиц. Механизмы активации фагоцитов обоих типов (полиморфноядерные лейкоциты и моноциты/макрофаги) принципиально одинаковы. Активирующими стимулами могут быть бактериальные продукты (например, ЛПС, N-формиловые пептиды и др.), компоненты комплемента (например, СЗ и С5), многие цитокины и AT, рецепторы к которым присутствуют на мембранах фагоцитов. Фагоцитоз состоит из четырёх последовательных стадий: хемотаксис, прикрепление к объекту, поглощение и уничтожение.
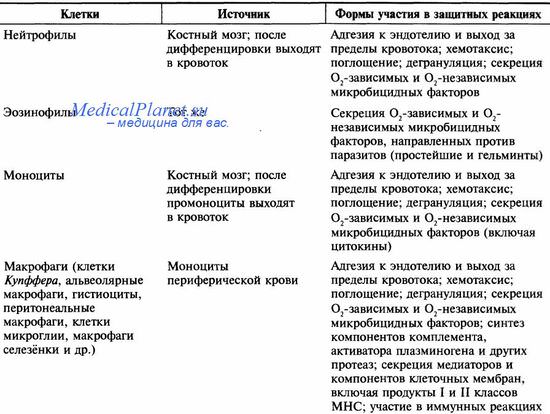
Хемотаксис — амебовидное передвижение фагоцитов по градиенту концентрации активирующих стимулов (хемотаксинов, или факторы хемотаксиса). Свойством активировать миграцию макрофагов обладают СЗb, С5а, C5b, C6-, С7-и Ва-компоненты комплемента, бактериальные липополисахариды, продукты деградации клеток, хемокины. Скорость привлечения клеток при хемотаксисе легко представить, оценив, например, время образования гноя и его объём после попадания занозы.
Адгезия. Одно из условий успешного поглощения возбудителя — эффективная адгезия к микробу. Жгутики позволяют микробам быстро перемещаться в жидкой фазе, а фагоциты не умеют «плавать», но хорошо «бегают», то есть свои поглотительные свойства они способны реализовывать только на какой-либо плотной поверхности (например, на эпителии). Опсонины, такие как AT, C3b, фибронектин, сурфактант, обволакивают микроорганизмы и существенно ограничивают их подвижность. Опсонины делают поглощение более эффективным, что связано со стабильностью взаимодействий опсонинов с соответствующими рецепторами (к Fc-фрагментам AT, компонентам комплемента, фибронектину и др.) на мембране фагоцита. Отсутствие этих рецепторов приводит к резкому снижению функциональной активности фагоцитов (например, врождённый дефицит С3b-рецепторов сопровождается высокой частотой бактериальных инфекций и даже выделен в отдельную нозологическую форму — недостаточность адгезии лейкоцитов).
Поглощение. Поглощение микробов идентично таковому у амёб; в результате образуется фагосома с заключённым внутри объектом фагоцитоза. К фагосоме устремляются лизосомы и выстраиваются по её периметру. Затем мембраны фагосомы и лизосом сливаются (фагосомо-лизосомальное слияние), и ферменты лизосом изливаются в образовавшуюся фаголизосому. Поглощению способствует взаимодействие поверхностных рецепторов фагоцитов с Аг или фрагментами опсонинов, сорбированных на поверхности бактерии. Эта реакция напоминает действие замка-молнии (так называемый зипперный механизм поглощения [от англ. zip, замок-молния]). Фагоцитированные микроорганизмы подвергаются атаке комплекса различных микробицидных факторов, разделяемых на кислородзависимые и кислороднезависимые.
Кислородзависимая микробицидная активность реализуется через образование значительного количества продуктов с токсическим действием, повреждающих микроорганизмы и окружающие структуры. За их образование ответственны НАДФ-оксидаза (флавопротеин цитохромредуктаза) плазматической мембраны и цитохром b; в присутствии хинонов этот комплекс трансформирует 02 в анион супероксида (02~). Последний проявляет выраженное повреждающее действие, а также быстро трансформируется в перекись водорода по схеме: 202~ + Н20 = Н202 + 02 (процесс катализирует фермент супероксид дисмутаза).
Пероксид водорода (Н202) проявляет меньший повреждающий эффект, но в его присутствии фермент миелопероксидаза конвертирует ионы С1~ в ионы НСЮ~, обладающие бактерицидным действием, во многом аналогичным эффекту хлорной извести (NaClO).
Кислороднезависимые механизмы активируются в результате контакта опсони-зированного объекта с мембраной фагоцита. В процессе фагосомолизосомального слияния первыми с мембраной фагосомы сливаются гранулы, содержащие лактоферрин и лизоцим, затем к ним присоединяются азурофиль-ные гранулы, содержащие катионные белки (например, САР57, САР37), протеиназы (например, эластазу и коллагеназу), катепсин G, дефензины и др. Эти продукты вызывают повреждение клеточной стенки и нарушение некоторых метаболических процессов; в большей степени их активность направлена против грамположительных бактерий.
Завершённость фагоцитарных реакций. Поглощённые фагоцитами бактерии обычно погибают и разрушаются. Некоторые бактерии, снабжённые капсулами или плотными гидрофобными клеточными стенками, могут быть устойчивы к действию лизосомальных ферментов; другая часть патогенов способна блокировать слияние фагосом и лизосом. В подобных случаях фагоцитоз носит незавершённый характер, и возбудитель выживает в цитоплазме поглотившей его клетки. Многие факультативные и облигатные внутриклеточные паразиты не только сохраняют жизнеспособность внутри клеток, но и способны размножаться. Персистирование патогенов опосредуют три основных механизма.
Блокада фагосомолизосомального слияния. Подобный механизм отмечен у вирусов (например, вируса гриппа), бактерий (например, микобактерий) и простейших (например, токсоплазм).
Резистентность к лизосомальным ферментам (например, гонококки и стафилококки).
Способность патогенных микроорганизмов быстро покидать фагосомы после поглощения и длительно пребывать в цитоплазме (например, риккетсии).
Другие защитные функции фагоцитов
• Инициация иммунных реакций. Поглощая чужеродные агенты, макрофаги «перерабатывают» (так называемый процессинг Аг) и «представляют» (презентация) их Аг иммунокомпетентным клеткам. При этом они выделяют цитокины, активирующие лимфоциты. Другими словами, макрофаги — Аг-процессирующие и Аг-представляющие клетки, и фагоцитоз можно рассматривать как видимое отражение этих иммунных функций.
• Антителозависимый цитолиз. Фагоциты участвуют в антителозависимом клеточно-опосредованном цитолизе. Для этого клетки экспрессируют на своей поверхности рецептор Fc-фрагмента IgG (CD 16).
- Читать далее "Интерфероны. Виды и функция интерферонов."
Оглавление темы "Система комплемента. Иммунодефициты.":1. Система комплемента. Классический путь системы комплемента.
2. Альтернативный путь системы комплемента. Фагоцитирующие клетки.
3. Интерфероны. Виды и функция интерферонов.
4. Естественные антитела. Естественные киллеры.
5. Иммунопатологические состояния. Причины, патогенез иммунопатологических состояний.
6. Иммунодефициты. Иммунодефицитные состояния.
7. Причины иммунодефицитов. Синдром ретикулярной дисгенезии.
8. Тяжёлый комбинированный иммунодефицит. Вариабельный общий иммунодефицит.
9. Синдром Незелофа. Синдром Иова. Лечение иммунодефицитов.
10. ВИЧ-инфекция. Синдром приобретённого иммунодефицита. СПИД.