
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Гладкая мышечная ткань: строение, гистология
Гладкая мышечная ткань состоит из удлиненных клеток, не обладающих поперечной исчерчен ностью, каждая из которых окружена базальной пластинкой и сетью ретикулярных волокон. Последние два компонента необходимы для того, чтобы объединять усилие, развиваемое отдельным гладким мышечным волокном, в совместное действие, например перистальтику в кишке.
Гладкие мышечные клетки имеют веретеновидную форму, т.е. их толщина максимальна в среднем участке, а к концам они конически сужаются. Длина этих клеток может колебаться от 20 мкм в мелких кровеносных сосудах до 500 мкм в матке при беременности. В матке во время беременности происходит значительное увеличение и размеров, и количества гладких мышечных клеток.
В каждой клетке имеется одно ядро, расположенное в центре наиболее широкой части клетки. С тем, чтобы в ткани достигалась наиболее плотная упаковка клеток, к узкой части одной клетки прилегают широкие части соседних клеток. Вследствие такого расположения на поперечном срезе ткани видны сечения клеток разнообразных диаметров, причем ядро обнаруживается только в наиболее крупных из них. При сокращении гладкой мышечной ткани границы клеток становятся фестончатыми, а ядро — складчатым или штопорообразным.
У полюсов ядра концентрируются митохондрии, полирибосомы, цистерны гранулярной эндоплазматической сети (грЭПС) и комплекс Гольджи. У клеточной поверхности часто обнаруживаются пиноцитозные пузырьки.
Саркоплазматическая сеть развита очень слабо; она состоит из замкнутой системы мембран, сходной с саркоплазматической сетью поперечнополосатой мышечной ткани. Т-трубочки в гладких мышечных клетках отсутствуют.
Характерная сократительная активность гладкой мышцы связана со структурой и организацией ее актиновых и миозиновых филаментов, которые не обладают паракристаллической организацией, имеющейся в поперечнополосатых мышцах. В гладких мышечных клетках пучки миофиламентов располагаются в цитоплазме под углом крест-накрест, образуя сеть. Эти пучки состоят из тонких филаментов (5— 7 нм), содержащих актин и тропомиозин, и толстых филаментов (12—16 нм), содержащих миозин.
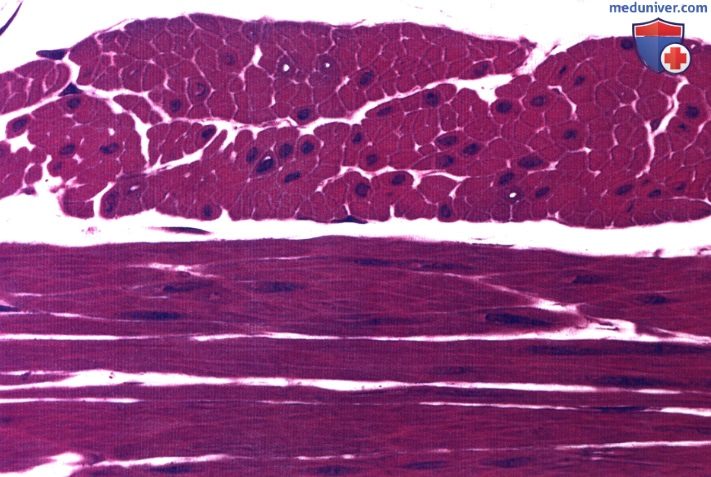
Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез. Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
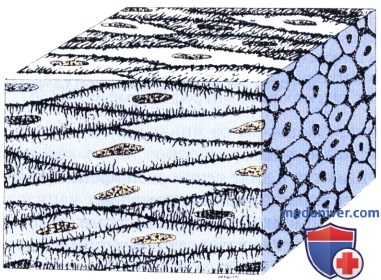
Как структурные, так и биохимические исследования показывают, что гладкомышечный актин и миозин взаимодействуют, обеспечивая сокращение посредством механизма скользящих филаментов, сходного с имеющимся в поперечнополосатых мышцах.
В гладких мышечных клетках сокращение начинается с притока Са2+. Миозин гладкой мышцы, однако, взаимодействует с актином, только когда его легкая цепь фосфорилирована. По этой причине, а также потому что тропомиозиновый комплекс, имеющийся в скелетной мышечной ткани, отсутствует механизм сокращения в гладкой мышечной ткани несколько отличается от такового в скелетной и сердечной мышечных тканях.
В гладких мышечных клетках присутствует кальмодулин — кальций-связывающий белок, который участвует также и в сокращении немышечных клеток. Са2+ образует комплексы с кальмодулином, которые активируют киназу легких цепей миозина — фермент, ответственный за фосфорилирование миозина.
Помимо кальция, на активность киназы легких цепей миозина влияют другие факторы, тем самым они воздействуют на степень сокращения гладких мышечных клеток. Сокращение или расслабление может регулироваться гормонами, действие которых опосредуется цАМФ. При повышении уровней цАМФ активируется киназа легких цепей миозина, происходит фосфорилирование миозина, и клетка сокращается. Снижение концентрации цАМФ оказывает обратное действие, уменьшая сократимость. Другим примером регуляции, не связанной с нервной системой, является действие половых гормонов на гладкую мышечную ткань матки.
Эстрогены вызывают увеличение концентрации цАМФ, обеспечивая фосфорилирование миозина и сократительную активность гладких мышечных клеток матки. Прогестерон дает противоположный эффект: он снижает уровни цАМФ, вызывая дефосфорилирование миозина и расслабление мускулатуры матки.
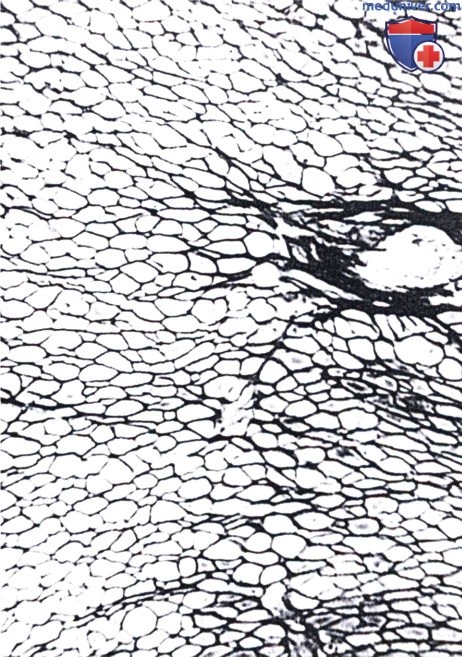
Эти волокна образуют сеть, окружающую мышечные клетки, которые данным методом не окрашиваются. Справа располагается артериола, окруженная толстыми коллагеновыми волокнами, х300.
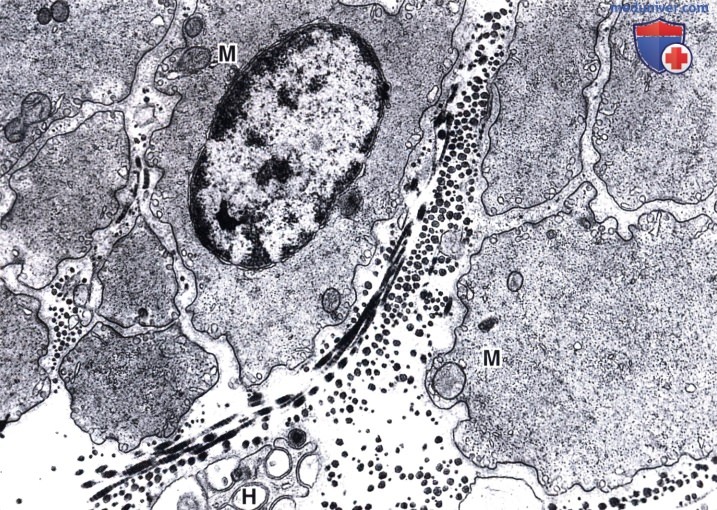
Толстые и тонкие филаменты не организованы в миофибриллы, митохондрии (М) немногочисленны. Обратите внимание на коллагеновые фибриллы, образующие ретикулярные волокна, и мелкое безмиелиновое нервное волокно (Н) между клеток.
Электронная микрофотография, х6650.
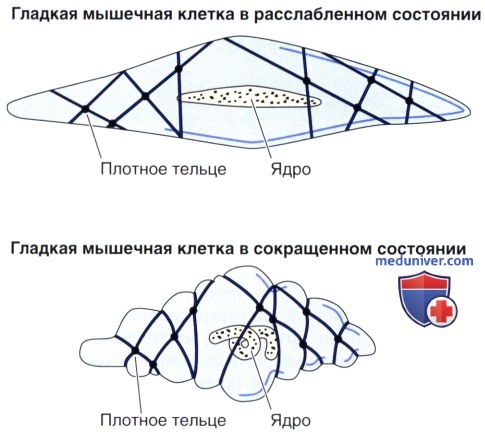
Сокращение этих филаментов вызывает укорочение клетки и обеспечивает сокращение всей мышцы. При сокращении ядро клетки деформируется.
Гладкие мышечные клетки содержат развитую систему промежуточных филаментов толщиной 10 нм, которые пронизывают их цитоплазму. Десмин (скелетин) идентифицирован как главный белок промежуточных филаментов во всех гладких мышечных клетках, дополнительным компонентом в сосудистой гладкой мышце является виментин. В гладких мышечных клетках выявляются плотные тельца двух типов. Один связан с мембраной; другой располагается в цитоплазме. Оба типа содержат а-актинин, тем самым они сходны с Z-линиями поперечнополосатой мышечной ткани.
Как тонкие, так и промежуточные филаменты внедряются в плотные тельца, которые передают сократительное усилие на соседние гладкие мышечные клетки и окружающую их сеть ретикулярных волокон.
Степень иннервации в определенном пучке гладкой мышечной ткани зависит от функции и размеров мышцы. Гладкие мышцы иннервируются как симпатическими, так и парасимпатическими нервами автономной (вегетативной) нервной системы. Развитые нервно-мышечные соединения, подобные тем, что имеются в скелетной мышце, в гладкой мышце отсутствуют. Аксоны автономных нервов частооканчиваются в виде серии расширенных участков в соединительной ткани эндомизия.
В целом гладкая мышечная ткань встречается в виде крупных пластов, например, в стенках полых внутренних органов — кишки, матки и мочеточников. Ее клетки обладают многочисленными щелевыми соединениями и относительно слабой иннервацией. Такие мышцы — висцеральные гладкие мышцы — функционируют как синцитий. Напротив, гладкие мышцы с множественными двигательными единицами обладают богатой иннервацией и могут осуществлять точные и регулируемые сокращения, такие, как, например, в радужке глаза.
Гладкая мышца обычно обладает спонтанной активностью в отсутствие нервной стимуляции. Поэтому функция ее иннервации заключается в модификации активности, а не в ее запуске, как в скелетной мышце. В гладкой мышце находятся как адренергические, так и холинергические нервные окончания, которые обладают антагонистическими эффектами, стимулируя или угнетая ее деятельность. В некоторых органах холинергические нервные окончания оказывают активирующее действие, а адренергические — угнетающее; в других их влияние имеет обратную направленность.
Помимо сократительной активности, гладкие мышечные клетки способны синтезировать коллаген, эластин и протеогликаны — компоненты межклеточного вещества, выработку которых обычно связывают с деятельностью фибробластов.
Видео лекция гистология мышечной ткани (гладкой, поперечнополосатой, сердечной)
- Читать далее "Регенерация мышечной ткани. Механизмы заживления мышц"
Оглавление темы "Гистология мышечной ткани":- Виды мышечной ткани. Строение
- Строение скелетной мышцы. Гистология
- Саркоплазматическая сеть и система поперечных трубочек скелетной мышечной ткани
- Механизмы сокращения скелетной мышцы. Схема
- Иннервация скелетных мышц. Механизмы
- Мышечные веретена и сухожильные органы Гольджи. Гистология
- Сердечная мышца: строение, гистология
- Гладкая мышечная ткань: строение, гистология
- Регенерация мышечной ткани. Механизмы заживления мышц