
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Генная конверсия. Механизмы генной конверсии.
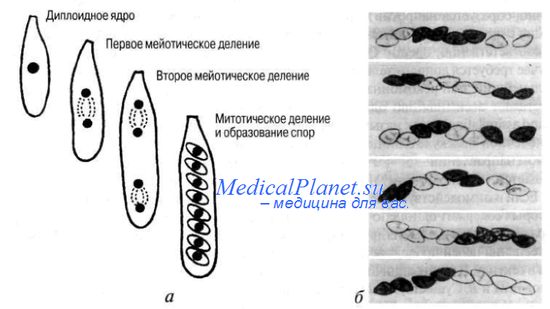
Генная конверсия — это процесс нереципрокного переноса информации из одной хроматиды в другую. Очень хорошим модельным объектом для изучения этого феномена служат некоторые виды низших грибов, например, Ascomycetes, продукты мейоза которых остаются в одной крупной клетке-сумке, называемой аском, располагаясь в линейном порядке. Если мы наблюдаем за гетерозиготным штаммом (Аа), то следовало бы ожидать распределение аллелей в 8 спорах аска 4А:4а, рекомбинация может лишь поменять местами споры с разными аллелями.
В 1928 г. Г. Книп обнаружил у нейроспоры отклонение от ожидаемого расщепления. Гетерозиготы Аа иногда давали споры в соотношении 3А : 1а или 1A : 3а и более редко-только А или только а. Такие же явления были отмечены в 1949-1953 гг. К. Линдегреном в опытах на дрожжах.
Г. Винклер, предложивший в 1930 г. назвать этот феномен конверсией, полагал, что в основе его - наличие у гетерозигот наследуемых изменений (мутаций) направленного характера, при которых переход одного аллеля в другой якобы инициируется гетерозиготным состоянием локуса. Но позже было установлено, что образование аберрантных асков не связано непосредственно с возникновением мутаций. А в 1955 г. М. Митчел обнаружила связь конверсии с кроссинювером и показала, что внутри генная рекомбинация может быть нереципрокной. Обнаружение аберрантных асков с соотношением спор 3:5 или 2:6 указывает на то, что мейотическому делению предшествовало возникновение гетеродуплексных участков кроссинговера между гомологичными материнской и отцовской хромосомами.
Как выяснилось при последующем изучении, протяженность этих участков может достигать 1 т.п.н.
Когда сочлененные участки гомологичных хромосом несколько различаются, могут произойти нарушения спаривания нуклеотидов, которые не останутся «незамеченными». Они будут исправлены системой репарации, предполагающей «сверку» ДНК и изменение одного из фрагментов ДНК по матрице другого. Под влиянием такой коррекции аллель В «превращается» в аллель в. Таким образом, изменение соотношения количества спор с разными аллелями в одном аске означает, что один из аллелей перешел в альтернативное состояние.
Если первоначально конверсия гена рассматривалась только применительно к нарушению классического менделевского расщепления у Ascomycetes, то в настоящее время этот термин имеет гораздо более широкое значение: его распространили на все процессы, включающие превращение одного аллеля в другой путем коррекции рекомбинационного гетеродуплекса. Причем установлено, что конверсия (нереципрокная гомологичная рекомбинация за счет коррекции неспаренных оснований в гетеродуплексе) происходит не только у прокариот, но и в половых, и в соматических клетках эукариот. В коррекции рекомбинационного гетеродуплекса у Е. coli используется описанный в предыдущей главе механизм репарации с участием генов системы MutHSL U. Для части бактериальных белков этой системы выявлены гомологи у эукариот (MHSLH - MutHSL homologs, или PMS — post meiotic segregation).
Причем, у эукариот гомологи представлены целыми семействами, но не все их члены заняты в конверсии. Так из шести гомологов MutS у дрожжей в конверсии участвуют четыре: MSHI, MSH2, MSH3 и MSH6. У человека обнаружено 11 гомологов MutL, из них участие в конверсии установлено для одного (PMS2). Показано, что конверсия у прокариот затрагивает небольшие фрагменты ДН К (не более трех нуклеотидов). В случае постмейотической сегрегации у эукариот изменяется несколько большая часть гена: от восьми до двенадцати нуклеотидов. Конверсия может иметь место и в случае митотической рекомбинации, но значительно реже. К общим закономерностям процесса конверсии следует отнести: 1) совместное конвертирование всех гетерозиготных маркеров, попавших в гетеродуплекс; 2) случайный выбор удаляемого основания из двух неспаренных; 3) полярность - снижение частот конверсии разных аллелей одного гена, начиная с определенной точки (чаще всего от одного конца гена к другому). Полярность указывает на наличие в хромосомах фиксированных точек начала рекомбинации.
- Читать далее "Сайт специфическая рекомбинация. Особенности сайт специфической рекомбинации."
Оглавление темы "Рекомбинация и транскрипция ДНК.":1. Атаксия-телеангиэктазия или синдром Луи-Бар. Синдром Блума.
2. Синдром Хатчинсона-Гилфорда или прогерия детей. Синдром Вернера.
3. Комбинативная изменчивость. Генетическая рекомбинация.
4. Модель Холлидея. Структура Холлидея.
5. Модель Мезельсона-Реддинга. Модель Жостака в генетике.
6. Генная конверсия. Механизмы генной конверсии.
7. Сайт специфическая рекомбинация. Особенности сайт специфической рекомбинации.
8. Незаконная рекомбинация в генетике. Методика незаконной рекомбинации.
9. Регуляция генной активности в ДНК. Транскрипция и регуляция генной активности.
10. Этапы транскрипции. Особенности транскрипции.