
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Группы сцепления генов в хромосоме. Понятие группы сцепления.
Число признаков, наследование которых соответствует менделевским закономерностям, много больше, чем число пар хромосом. Следовательно, в одной хромосоме должно находиться много генов, контролирующих разные признаки. Гены, расположенные в одной хромосоме, должны чаще наследоваться вместе, т.е. для них должен нарушаться закон Менделя о независимом наследовании. На заре развития генетики это был аргумент против хромосомной гипотезы наследственности. Однако по мере накопления данных становилось все более очевидным, что закон независимого наследования действительно соблюдается не для любых пар признаков. Феномен сцепления впервые установили в 1906 г. У. Бэтсон и Р. Пеннет, изучая наследование некоторых признаков у душистого горошка.
Они обнаружили, что у гибридов фактор, определяющий развитие удлиненных пыльцевых зерен (L), чаще попадает в те гаметы, куда попал и фактор фиолетовой окраски (В). Такое совместное наследование они назвали гаметинеским сцеплением. Для других признаков, например, для окраски цветка и формы паруса, наоборот, было обнаружено, что фактор фиолетовой окраски (В) и фактор прямого паруса (Е) редко попадают в одну гамету. Бэтсон предположил, что в этом случае происходит «гаметическое отталкивание» факторов друг от друга. Явления притяжения и отталкивания Бэтсон и Пеннет объяснили увеличением числа гамет, несущих определенные сочетания наследственных факторов. Они никак не связали эти явления с хромосомной гипотезой наследственности.
Классическими исследованиями сцепленного наследования явились работы Т Моргана и его сотрудников на дрозофиле. Довольно быстро у дрозофилы было обнаружено и проанализировано наследование сотен мутаций, изменяющих разнообразные признаки. Все гены можно было разделить на четыре группы, которые назвали группами сцепления. Число групп сцепления равно числу пар хромосом. Одна группа включала гены, контролирующие признаки, сцепленные с полом, две большие группы и одна малочисленная — гены, контролирующие признаки, наследующиеся по менделевским правилам. Гены, относящиеся к одной группе, наследуются сцепленно, т.е. потомки чаще несут родительские комбинации признаков. Гены, принадлежащие к разным группам, наследуются независимо.
У дрозофилы четыре пары хромосом и четыре группы сцепления, причем число генов, обнаруженных для каждой группы сцепления, коррелировало с длиной хромосомы.
Тот факт, что расположение генов в одной хромосоме приводит к нарушению закономерностей независимого наследования, можно показать для генов, локализованных в Х-хромосоме. Рассмотрим опыты А. Стертеванта по изучению совместного наследования признаков «белые глаза» (ген w) и «желтое тело» (ген у). Легко показать, что гены, определяющие эти признаки, расположены в Х-хромосоме. Если самок у w/y w, гомозиготных по рецессивным генам, скрестить с самцами дикого типа, у+ w+/Y в потомстве появятся самцы с фенотипом у w (желтое тело и белые глаза) и гетерозиготные самки дикого типа. Скрещивание самок и самцов F1 является анализирующим, оно позволяет установить частоты гамет разных типов, образуемых гетерозиготными самками. В F2 было получено:
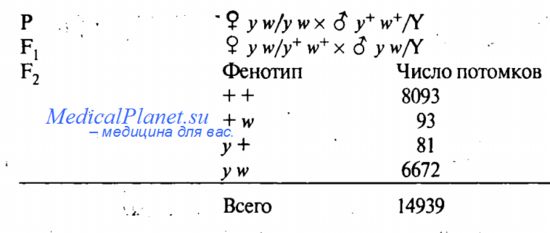
Подавляющее большинство потомков (+ + и у w) имеет родительские комбинации генов. Число потомков + w и у +, с новыми комбинациями генов (рекомбинантные потомки) составляет всего 1,2%. Итак, новые комбинации появились, но не в том соотношении, которое ожидается при независимом наследовании признаков (1:1:1:1).
Скрещивание желтых самок с белоглазыми самцами в F1 дает гетерозиготных самок, но в отличие от предыдущего случая мутации у и w находятся у них в транс-положении, т.е. в разных хромосомах. Анализирующее скрещивание дало следующие результаты:

Вновь родительские комбинации признаков (+ w и у +) обнаруживаются у большинства потомков, а рекомбинантные (+ + и у w) - только у 1,4%. Доли рекомбинантных потомков в двух экспериментах практически равны. Показано, что для каждой пары сцепленных генов характерна определенная доля рекомбинантных потомков, не зависящая от направления скрещивания и родительских комбинаций генов. Для разных генов частота рекомбинации варьирует от 0 до 50%. На основании изучения частот рекомбинации между сцепленными генами строят генетические карты хромосом.
- Читать далее "Теория наследственности Моргана. Аутосомно-доминантное наследование признаков."
Оглавление темы "Хромосомы и гены.":1. Хромосомы. Хромосомная теория наследственности.
2. Клеточный цикл. Хромосомы в ходе клеточного цикла. Митоз и фазы митоза.
3. Мейоз. Фазы мейоза. Процессы во время мейоза.
4. Половые различия в типе хромосом. Признаки сцепленные с полом.
5. Нерасхождение половых хромосом. Изучение нерасхождения половых хромосом.
6. Группы сцепления генов в хромосоме. Понятие группы сцепления.
7. Теория наследственности Моргана. Аутосомно-доминантное наследование признаков.
8. Аутосомно-рецессивный тип наследования признаков. Недоминантное наследование.
9. Ген. История изучения генов. Роль гена в мутации и наследственности.
10. Молекула ДНК. Открытие ДНК. Генетическая роль ДНК.