
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизмы восстановления (регенерации) печени
Печень человека имеет значительную способность к регенерации, о чем свидетельствует ее рост после частичной гепатэктомии, которую выполняют по поводу резекции опухоли или при трансплантации печени от живого донора. Известным примером регенерации печени является восстановление печени Прометея, которую каждый день выклевывал орел, присланный Зевсом (Зевс разгневался на Прометея за кражу огня, но знал ли он, что печень Прометея будет регенерировать?).
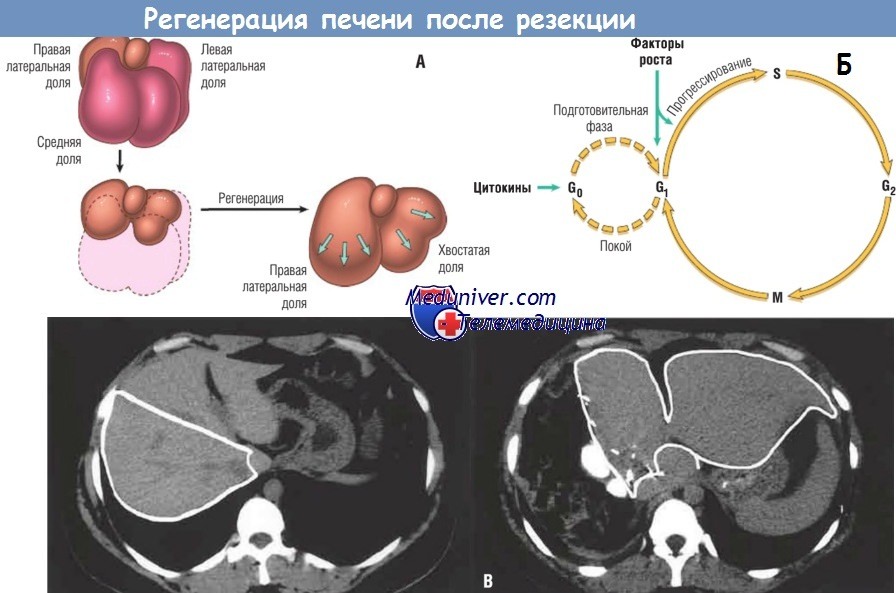
Реальность, хотя и не столь сказочная, все равно впечатляет. Резекция около 60% печени у живого донора приводит к удвоению остатка печени в течение примерно 1 мес. Часть органа, оставшаяся у донора после резекции, является «мини-печенью», которая быстро увеличивается и достигает исходной массы. Восстановление массы печени происходит не за счет повторного роста долей, а путем увеличения оставшихся после операции долей. Данный процесс называют компенсаторным ростом или компенсаторной гиперплазией. Как у людей, так и у грызунов конечным результатом регенерации печени после частичной резекции является восстановление функциональной массы, а не воссоздание оригинальной структуры.
При регенерации печени после частичной резекции почти все гепатоциты начинают делиться. Поскольку гепатоциты являются покоящимися клетками, вход в клеточный цикл, прохождение фазы G1 и достижение фазы S репликации ДНК занимает у них несколько часов. Подъем репликации гепатоцитов синхронизирован и сопровождается синхронной репликацией непаренхиматозных клеток (клеток Купфера, эндотелиальных и звездчатых клеток).
Существует немало доказательств того, что пролиферация гепатоцитов в регенерирующей печени вызвана совместным действием цитокинов и полипептидных факторов роста. За исключением аутокринного действия TGF-a, репликация гепатоцитов строго зависит от паракринных эффектов факторов роста и цитокинов — HGF и IL-6, продуцируемых непаренхиматозными клетками печени. Существует две основные точки ограничения в репликации гепатоцитов: в переходе G0/G1, когда покоящиеся гепатоциты вступают в клеточный цикл, и в переходе G1/S, необходимом для прохождения в конце фазы G1 точки ограничения.
Экспрессия генов в регенерирующей печени протекает по фазам, начиная с немедленного раннего генного ответа, который является временным и соответствует переходу G0/G1. В этот период активируются более 70 генов, в т.ч. протоонкогены c-FOS и c-JUN, продукты которых димеризуются с образованием фактора транскрипции АР-1, и ген c-MYC, который кодирует активирующий различные гены фактор транскрипции, а также другие факторы транскрипции, например NF-kB, STAT-3 и С/ЕВР. Немедленный ранний генный ответ создает условия для последовательной активации многочисленных генов во время перехода гепатоцитов в фазу G1. Переход из фазы G1 в фазу S описан ранее.
Покоящиеся гепатоциты приобретают способность вступать в клеточный цикл через подготовительную фазу, в основном при участии цитокинов TNF и IL-6 и компонентов системы комплемента. Своим участием они активируют несколько сигнальных путей в качестве необходимой подготовки к клеточной пролиферации. Стимулируемые HGF, TGF-a и гепаринсвязывающим EGF-подобным фактором роста покоящиеся гепатоциты вступают в клеточный цикл, и происходит репликация ДНК. Норадреналин, серотонин, инсулин, тиреоидный гормон и гормон роста при регенерации печени действуют в качестве добавочных факторов, способствуя вступлению гепатоцитов в клеточный цикл.
В процессе регенерации отдельные гепатоциты делятся 1 или 2 раза, а затем возвращаются в состояние покоя, строго следуя определенной последовательности событий. Механизмы прекращения репликации еще не установлены. В прекращение репликации гепатоцитов могут быть вовлечены ингибиторы роста, например TGF-b и активины, но механизм их действия еще не ясен. В компенсаторном росте органа после частичной гепатэктомии принимают участие внутрипеченочные стволовые клетки, или клетки-предшественники, но в данной ситуации нет доказательств образования гепатоцитов из клеток костного мозга. Однако эндотелиальные и другие непаренхиматозные клетки регенерирующей печени могут происходить от клеток-предшественников из костного мозга.
Показаны рецепторы, обладающие собственной тирозинкиназной активностью, 7-трансмембранные G-белок-связанные рецепторы и рецепторы, не обладающие собственной тирозинкиназной активностью.
1Р3 — инозитол-1,4,5-трифосфат; JAK — Janus-киназы; МАРК — митоген-активируемая протеинкиназа;
PI3K — фосфатидилинозитол-3-киназа; РКВ — протеинкиназа В, также известная как Akt;
PLC-y— фосфолипаза С-7; STAT — трансдукторы сигнала и активаторы транскрипции;
ДНК — дезоксирибонуклеиновая кислота; цАМФ — циклический аденозинмонофосфат.
- Рекомендуем ознакомиться со следующей статьей "Взаимодействие межклеточного вещества (внеклеточного матрикса, ВКМ) с клеткой"
Оглавление темы "Патофизиология клетки и тканей":- Функции стволовых клеток в тканях и органах
- Фазы клеточного цикла и клеточная репликация
- Факторы роста клеток и тканей
- Механизмы передачи сигнала роста между клетками
- Рецепторы и сигнальные пути клеток
- Факторы транскрипции клетки - функции, значение
- Механизмы восстановления (регенерации) органов и тканей
- Механизмы восстановления (регенерации) печени
- Взаимодействие межклеточного вещества (внеклеточного матрикса, ВКМ) с клеткой
- Типы коллагена межклеточного вещества (внеклеточного матрикса)