
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Внутренний путь развития апоптоза
Внутренний (митохондриальный) путь развития апоптоза — основной механизм его развития во всех клетках млекопитающих. Его роль в различных физиологических и патологических процессах хорошо изучена. Этот путь развития апоптоза — результат повышения митохондриальной проницаемости и высвобождения проапоптозных молекул (индукторов смерти клетки) в цитоплазму.
Митохондрии — основные органеллы клетки, содержащие такие белки, как цитохром С, которые жизненно необходимы, но некоторые из них при высвобождении в цитоплазму (признак нездоровья клетки) инициируют апоптоз. Высвобождение этих митохондриальных белков четко контролируется балансом между про- и антиапоптозными белками семейства Bcl.
Название семейства образовано от Bcl-2, онкогена в В-клеточных лимфомах, гомологичного белку Ced-9 С. elegans. Семейство Bcl включает более 20 белков, большинство из них регулируют апоптоз. Факторы роста и другие факторы выживания стимулируют продукцию антиапоптозных белков, основными среди них являются Bcl-2, Bcl-х и Мсl-1.
В норме они располагаются в цитоплазме на мембране митохондрии, где контролируют проницаемость ее мембраны и предотвращают выход митохондриальных белков, способных вызвать смерть клетки. Когда клетки лишаются сигналов к выживанию либо повреждается их ДНК или неправильно свернутые белки индуцируют ЭПР-стресс, активируются сенсоры повреждения.
Эти сенсоры также принадлежат к семейству Bcl и представлены белками Bim, Bid и Bad, которые содержат одиночный Bcl-2-гомологичный домен (третий из четырех таких доменов, присутствующих в Bcl-2). Эти белки называют белками ВН3-only. В свою очередь, сенсоры активируют два основных (проапоптозных) эффектора Вах и Bak, формирующих олигомеры, которые встраиваются в мембрану митохондрии и образуют каналы, позволяющие белкам внутренней мембраны митохондрии просочиться в цитоплазму.
Белки ВН3-only могут связаться с Bcl-2 и Bcl-х и блокировать их функцию. В то же время может снизиться синтез Bcl-2 и Bcl-х. Конечный результат активации Вах/Вак вместе с утратой защитных функций антиапоптозных белков семейства Bcl — это высвобождение в цитоплазму нескольких митохондриальных белков, активирующих каспазы. Одним из них является цитохром С, играющий важную роль в дыхании митохондрий.
При высвобождении в цитозоль цитохром С связывается с белком APAF1 (фактор активации протеаз апоптоза 1, гомологичный Ced-4 С. elegans), который формирует подобный колесу гексамер, названный апоптосомой. Этот комплекс способен связывать каспазу-9, основной инициатор внутреннего пути развития апоптоза, и соседние молекулы расщепляющих ферментов, тем самым начиная процесс аутоамплификации.
Другие митохондриальные белки — Smac/DIABLO — входят в цитоплазму, где присоединяются к нейтрализованным цитоплазматическим белкам, являющимся физиологическими ингибиторами апоптоза. Нормальной функцией ингибиторов апоптоза является блокировка активации каспаз, включая эффекторную каспазу-3, и поддержание клетки в живом состоянии. Таким образом, нейтрализация ингибиторов апоптоза позволяет инициировать каспазный каскад.
Существует ряд доказательств, что внутренний путь развития апоптоза может быть активирован и без участия митохондрий. Апоптоз может начаться в результате активации каспазами верхнего потока митохондрий, и последующие повышение митохондриальной проницаемости и высвобождение проапоптозных молекул усиливают сигнал к запуску апоптоза.
Механизмы развития апоптоза с независимой от митохондрий инициацией до конца не изучены.
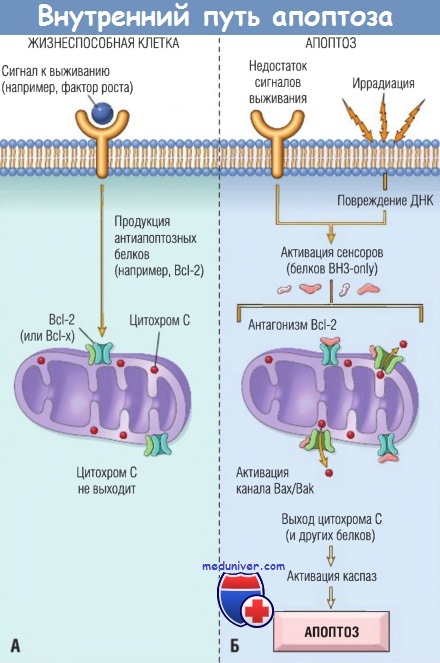
(А) Жизнеспособность клеток поддерживается индукцией сигналов к выживанию антиапоптозных белков, например Bcl-2.
Эти белки обеспечивают целостность мембран митохондрий и предотвращают выход митохондриальных белков.
(Б) Потеря сигналов к выживанию, повреждение дезоксирибонуклеиновой кислоты (ДНК) и другие факторы, активирующие сенсоры,
которые противодействуют антиапоптозным белкам и активируют проапоптозные белки Вах и Bak, формирующие каналы в мембране митохондрий.
Последующий выход цитохрома С (и других белков; не показаны) ведет к активации каспаз и апоптозу.
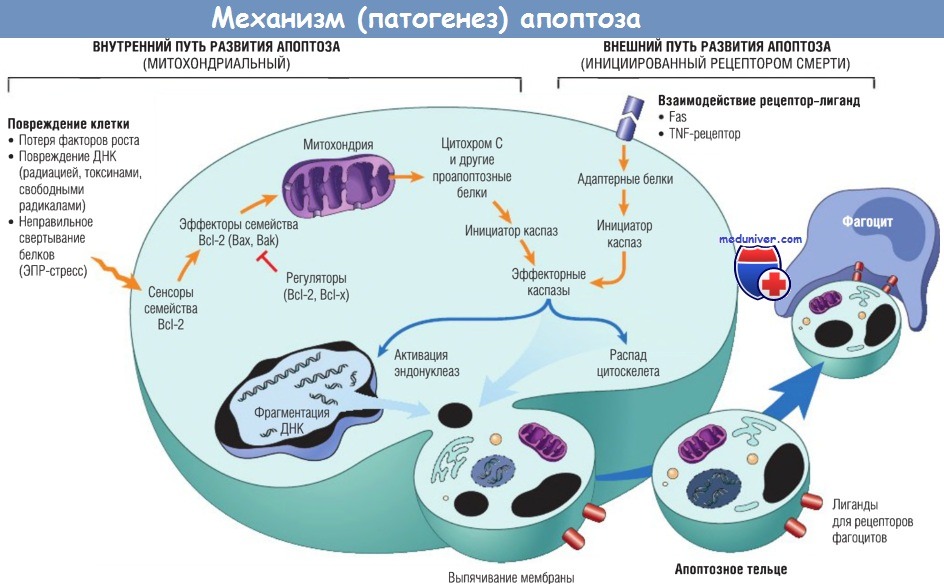
Два пути развития апоптоза отличаются индукцией и регуляцией, но оба заканчиваются активацией эффекторных каспаз.
При внутреннем пути развития апоптоза индукция вовлекает сенсоры и эффекторы семейства Bcl-2, которые индуцируют выход митохондриальных белков.
Показаны также некоторые антиапоптозные белки (регуляторы), которые ингибируют митохондриальный выход и активацию цитохром С-зависимой каспазы во внутреннем пути.
При внешнем пути развития апоптоза инициация рецепторов смерти напрямую вызывает активацию каспаз.
Регуляторы активации каспаз, опосредованные рецептором смерти, не показаны.
TNF — фактор некроза опухоли; ДНК — дезоксирибонуклеиновая кислота; ЭПР — эндоплазматический ретикулум.
- Рекомендуем ознакомиться со следующей статьей "Внешний путь развития апоптоза"
Оглавление темы "Патология клетки":- Ишемическое и гипоксическое повреждение клеток
- Реперфузионное повреждение клетки после ишемии
- Механизмы химического повреждения клетки
- Что такое апоптоз? Причины апоптоза
- Морфологические изменения при апоптозе
- Биохимические признаки апоптоза
- Механизмы апоптоза и их патогенез
- Внутренний путь развития апоптоза
- Внешний путь развития апоптоза
- Механизмы запуска апоптоза