
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизмы гиперчувствительности немедленного типа (I типа)
Гиперчувствительность типа I (немедленного типа) представляет собой быстро развивающуюся иммунологическую реакцию (в течение нескольких минут) после взаимодеиствия антигена с антителами, связанными с тучными клетками, у индивидов, сенсибилизированных антигеном. Эти реакции часто называют аллергией, а антигены, вызывающие ее, — аллергенами. Гиперчувствительность типа I может проявляться как системное расстройство или быть местной реакцией.
Системное расстройство обычно возникает вслед за инъекцией антигена сенсибилизированному индивиду. Иногда в течение нескольких минут пациент впадает в состояние шока, который может быть фатальным. Местные реакции разнообразны и варьируют в зависимости от входных ворот антигена: от локальной кожной припухлости (кожная аллергия, крапивница), выделений из носа и конъюнктивы (аллергические ринит и конъюнктивит) до сенной лихорадки, бронхиальной астмы или аллергического гастроэнтерита (пищевая аллергия).
Многие местные реакции гиперчувствительности типа I имеют две хорошо выраженные фазы. Немедленная, или инициальная, фаза характеризуется вазодилатацией, повышением сосудистой проницаемости и, в зависимости от локализации, спазмом гладких мышц или секрецией слизи. Эти явления обычно наблюдаются через 5-30 мин после воздействия аллергена и, как правило, уменьшаются через 60 мин.
Однако во многих случаях (например, при аллергическом рините и бронхиальной астме) через 2-24 час наступает поздняя фаза без дополнительного воздействия антигена, которая может длиться несколько суток. Эта фаза характеризуется инфильтрацией тканей эозинофилами, нейтрофилами, базофилами, моноцитами и Т-клетками CD4+, а также разрушением тканей (обычно повреждением эпителиальных клеток слизистых оболочек).
Большинство реакций гиперчувствительности типа I опосредовано зависимой от IgE-антител активацией тучных клеток и других лейкоцитов. Поскольку тучные клетки играют центральную роль в развитии гиперчувствительности типа I, следует сначала охарактеризовать некоторые их важные свойства. Тучные клетки имеют костномозговое происхождение и широко распространены в тканях. Большое количество тучных клеток присутствует вблизи кровеносных сосудов, нервов и в субэпителиальных тканях; это объясняет, почему местные реакции гиперчувствительности типа I часто наблюдаются в этих местах.
Тучные клетки содержат мембраносвязанные цитоплазматические гранулы, в которых находятся разнообразные биологически активные медиаторы. Гранулы также содержат кислые протеогликаны, связывающие основные красители, например толуидиновый синий. Тучные клетки (и базофилы) активируются в результате перекрестного связывания высокоаффинных Fc-рецепторов IgE. Кроме того, тучные клетки могут активировать такие стимулы, как компоненты системы комплемента С5а и С3а (называемые анафилатоксинами, т.к. они вызывают реакции, сходные с анафилаксией), действующие посредством связывания с рецепторами на мембране тучных клеток.
Секрецию тучных клеток повышают некоторые хемокины (в частности, IL-8), лекарственные средства (например, кодеин и морфин, аденозин, меллитин, присутствующий в пчелином яде) и физические факторы (тепло, холод, солнечный свет).
Базофилы во многом сходны с тучными клетками, включая присутствие Fc-рецепторов для IgE на поверхности клеток и цитоплазматических гранул. Однако в отличие от тучных клеток базофилы в обычных условиях в тканях отсутствуют, а циркулируют в кровотоке в крайне малом количестве. (Большинство аллергических реакций происходит в тканях, и роль базофилов в этих процессах изучена менее детально, чем роль тучных клеток.) Подобно другим гранулоцитам, базофилы могут мигрировать в очаг воспаления.
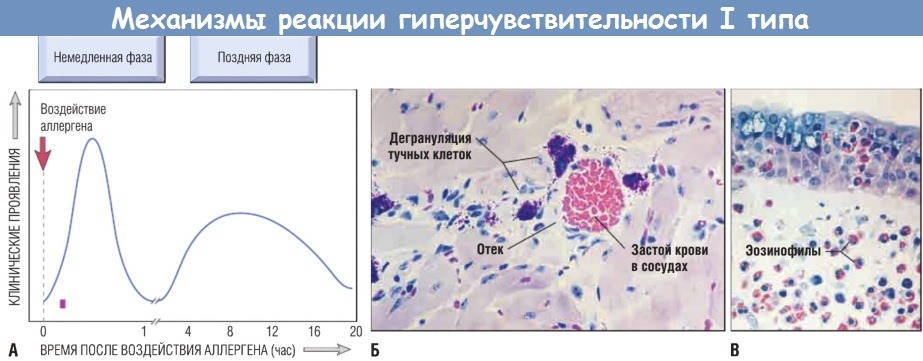
(А) Кинетика немедленной и поздней фаз реакции. Немедленная фаза — это реакция клеток гладких мышц и сосудов на аллерген через несколько минут после его воздействия на сенсибилизированного индивида; поздняя фаза развивается через 2-24 час.
(Б, В) Морфология немедленной фазы характеризуется расширением сосудов, застоем крови и отеком (Б);
морфология поздней фазы — воспалительным инфильтратом, богатым эозинофилами, нейтрофилами и Т-клетками (В)
Тh2-клетки играют главную роль в инициации и развитии реакций гиперчувствительности типа I, стимулируя продукцию IgE и способствуя воспалению. Первым этапом в генерации Тh2-клеток является презентация антигена хелперным Т-клеткам CD4+, вероятно, дендритными клетками, захватывающими антиген в месте его проникновения. В ответ на антиген и другие стимулы, включающие цитокины (например, IL-4), продуцируемые местно, Т-лимфоциты дифференцируются в Тh2-клетки. При последующей встрече с антигеном новообразованные Тh2-клетки секретируют ряд цитокинов.
Типичными для этой субпопуляции цитокинами являются IL-4, IL-5 и IL-13. IL-4 действует на В-клетки, стимулируя переключение класса на IgE и способствуя появлению дополнительных Тм2-клеток. IL-5 участвует в развитии и активации эозинофилов, которые, как будет показано далее, представляют собой важные эффекторы гиперчувствительности типа I. IL-13 усиливает продукцию IgE и действует на эпителиальные клетки, стимулируя секрецию слизи. Кроме того, Тh2-клетки (помимо тучных и эпителиальных клеток) продуцируют хемокины, привлекающие дополнительное количество Тh2-клеток и других лейкоцитов в место реакции.
Тучные клетки и базофилы экспрессируют высокоаффинный рецептор FceRI, специфический к Fc-фрагменту IgE и поэтому авидно связывающийся с IgE-антителами. Когда тучная клетка, несущая IgE-антитела, подвергается воздействию специфического аллергена, происходит серия реакций, приводящих к высвобождению целого арсенала высокоактивных медиаторов, ответственных за клинические проявления реакций гиперчувствительности типа I. На первом этапе антиген (аллерген) связывается с IgE-антителами, уже фиксированными на поверхности тучных клеток.
Поливалентные антигены связываются между собой и перекрестно связывают соседние IgE-антитела и, соответственно, Fc-рецепторы IgE. Такое перекрестное связывание Fcζ-рецепторов активирует пути передачи сигналов, посылаемых цитоплазматическим компонентом рецепторов. Эти сигналы вызывают дегрануляцию тучных клеток с высвобождением преформированных (первичных) медиаторов, хранившихся в гранулах, а также синтез de novo и высвобождение вторичных медиаторов, включая липидные продукты и цитокины. Эти медиаторы ответственны за инициальные, иногда возникающие внезапно симптомы гиперчувствительности типа I; они также инициируют процессы, приводящие к развитию реакции поздней фазы.
а) Преформированные медиаторы. Медиаторы, уже содержащиеся в гранулах тучных клеток, высвобождаются в первую очередь. Эти медиаторы можно подразделить на:
- вазоактивные амины. Наиболее важным из них является гистамин, который вызывает интенсивное сокращение гладких мышц, повышает сосудистую проницаемость и увеличивает секрецию слизи железами слизистой оболочки носа, бронхов и желудка;
- ферменты. Они содержатся в матриксе гранул и включают нейтральные протеазы (химазу, триптазу) и различные кислые гидролазы. Ферменты повреждают ткани и приводят к образованию кининов и активированных компонентов системы комплемента (например, С3а), действуя на белки-предшественники;
- протеогликаны. К ним относятся гепарин, хорошо известный антикоагулянт, и хондроитинсульфат. Протеогликаны нужны для «упаковки» и сохранения аминов в гранулах.
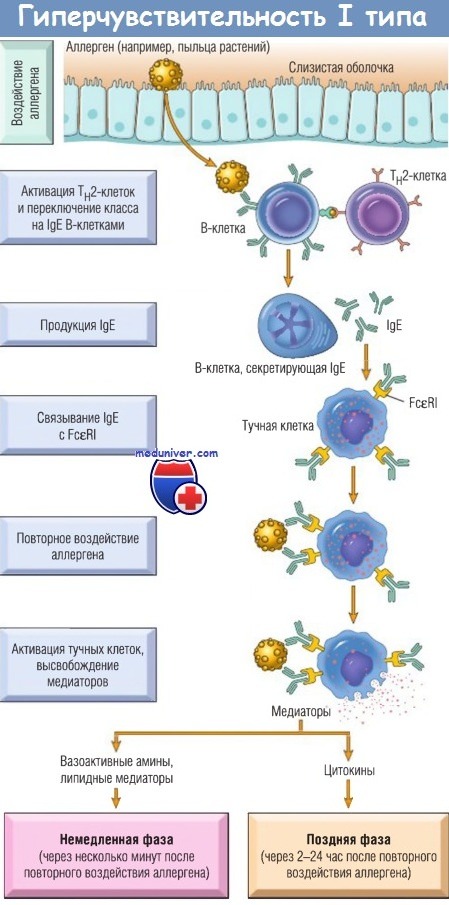
Реакцию гиперчувствительности типа I инициирует введение аллергена, стимулирующего ответ Тh2-клеток и продукцию имунноглобули-на Е (IgE) у генетически чувствительных индивидов.
IgE связываются с Fc-рецепторами (FceRI) на тучных клетках, и последующее воздействие аллергена активирует тучные клетки,
которые секретируют медиаторы, ответственные за патологические проявления немедленной и поздней фаз.
б) Липидные медиаторы. Основные липидные медиаторы синтезируются в результате последовательных реакций на мембранах тучных клеток. Эти реакции приводят к активации фосфолипазы А2 — фермента, ответственного за образование из фосфолипидов мембраны арахидоновой кислоты. Из этого исходного соединения под влиянием 5-липоксигеназы и циклооксигеназы образуются лейкотриены и простагландины:
- лейкотриены. Лейкотриены С4 и D4 являются наиболее сильнодействующими вазоактивными и спазмогенными агентами. По показателям повышения сосудистой проницаемости и сокращения гладких мышц бронхов они в несколько тысяч раз более активны, чем гистамин (в расчете на молярность). Лейкотриен В4 обладает выраженными хемотаксическими свойствами в отношении нейтрофилов, эозинофилов и моноцитов;
- простагландин D2. Этот медиатор образуется в тучных клетках в наибольшем количестве по циклооксигеназному пути и вызывает интенсивный бронхоспазм, а также повышенную секрецию слизи;
- PAF. Этот фактор образуют некоторые популяции тучных клеток. Он активирует агрегацию тромбоцитов, вызывает высвобождение гистамина, бронхоспазм, повышает сосудистую проницаемость и увеличивает вазо-дилатацию. Кроме того, он хемотаксичен для нейтрофилов и эозинофилов, а в больших концентрациях активирует клетки воспаления, вызывая их дегрануляцию. Хотя продукцию PAF также индуцирует фосфолипаза А2, он не является продуктом метаболизма арахидоновой кислоты.
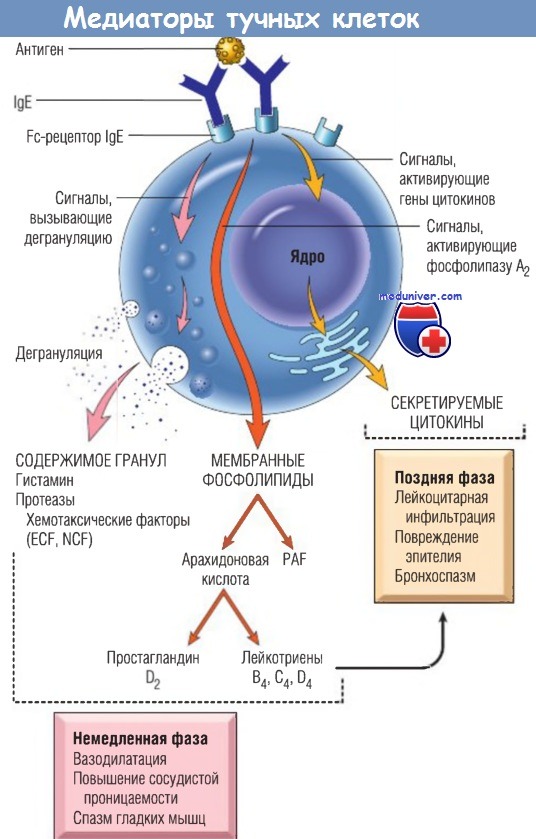
При активации тучные клетки высвобождают медиаторы различных классов, ответственные за немедленную фазу и позднюю фазу реакции.
ECF и NCF не определены биохимически.
ECF — эозинофильный хемотаксический фактор; Ig — иммуноглобулин;
NCF — нейтрофильный хемотаксический фактор; PAF — фактор активации тромбоцитов.
в) Цитокины. Тучные клетки служат источником многих цитокинов, которые играют важную роль на различных стадиях реакций гиперчувствительности типа I. К таким цитокинам относятся TNF, IL-1 и хемокины, способствующие рекрутированию лейкоцитов (типичный процесс для поздней фазы), IL-4, усиливающий ответ Тh2-клеток, и многие другие. Клетки воспаления, рекрутируемые TNF и хемокинами тучных клеток, представляют собой дополнительный источник цитокинов и высвобождающих гистамин факторов, обусловливающих дальнейшую дегрануляцию тучных клеток.
Развитие реакций гиперчувствительности типа I зависит от координированного действия разнообразных хемотаксических, вазоактивных и спазмогенных соединений. Некоторые из них, в частности гистамин и лейкотриены, быстро высвобождаются из сенсибилизированных тучных клеток и ответственны за интенсивную немедленную фазу, характеризующуюся отеком, секрецией слизи и спазмом гладких мышц. Другие, представленные цитокинами, обусловливают позднюю фазу в результате рекрутирования дополнительных лейкоцитов. Эти клетки воспаления не только высвобождают еще некоторое количество медиаторов (включая цитокины), но и приводит к повреждению эпителиальных клеток. Сами эпителиальные клетки также могут продуцировать медиаторы, в частности хемокины.
Среди клеток, мобилизуемых в позднюю фазу, особенно важны эозинофилы. Они рекрутируются в участки реакции гиперчувствительности типа I хемокинами (например, эотаксином), продуцируемыми эпителиальными клетками, Тh2-клетками и тучными клетками. Выживанию эозинофилов в тканях способствуют IL-3, IL-5 и гранулоцитарно-макрофагальный колониестимулирующий фактор. IL-5 является наиболее сильным активирующим эозинофилы цитокином из числа известных. Эозинофилы высвобождают протеолитические ферменты, а также два уникальных белка — главный основной белок и эозинофильный катионный белок, токсичные для эпителиальных клеток.
Активированные эозинофилы и другие лейкоциты также продуцируют лейкотриен С4 и PAF и непосредственно стимулируют высвобождение медиаторов тучными клетками. Таким образом, рекрутированные клетки амплифицируют и поддерживают воспалительную реакцию, не нуждаясь в дополнительном воздействии соответствующего антигена. В настоящее время считают, что поздняя фаза служит основной причиной симптомов при некоторых расстройствах, связанных с гиперчувствительностью типа I, например при аллергической бронхиальной астме. Для лечения этих заболеваний необходимо применение противовоспалительных лекарственных средств широкого спектра действия, в частности стероидов.
Предрасположенность к реакциям гиперчувствительности типа I генетически детерминирована. Термин атопия означает предрасположенность к развитию локализованных реакций гиперчувствительности типа I на разнообразные вдыхаемые или принимаемые с пищей антигены. У индивидов с атопией обнаруживаются более высокий уровень IgE в сыворотке и большее число IL-4-продуцирующих Тh2-клеток, чем в общей популяции. Такие индивиды в 50% случаев имеют аллергию в семейном анамнезе.
Основы наследственной предрасположенности неясны, однако исследования, проведенные с участием пациентов с астмой, указывают на сцепление с некоторыми генными локусами. Гены-кандидаты были картированы на хромосоме 5q31, где локализованы гены, кодирующие цитокины IL-3, IL-4, IL-5, IL-9, IL-13 и гранулоцитарно-макрофагальный колониестимулирующий фактор. Этот локус привлекает большое внимание, поскольку хорошо известна роль многих из этих цитокинов в реакции, однако непонятно, как влияет ассоциированный с заболеванием полиморфизм на биологию цитокинов.
Отмечено также сцепление с 6р в непосредственной близости к комплексу HLA, указывающее на то, что наследование определенных аллелей HLA определяет реактивность к некоторым аллергенам.
Значительная часть реакций гиперчувствительности типа I возникает в результате воздействия экстремальных температур и физических нагрузок, без участия Тh2-клеток и IgE-антител. Иногда такие реакции называют неатопической аллергией. Предполагается, что в подобных случаях тучные клетки обладают аномальной чувствительностью к активации различными неиммунными стимулами.
Последнее, о чем необходимо упомянуть при обсуждении общих вопросов патологии, связанных с гиперчувствительностью типа I, — это повышение частоты таких расстройств в развитых странах. Возможно, это связано со снижением частоты инфекционных заболеваний в ранний период жизни. Подобные наблюдения привели к предположению, называемому гигиенической гипотезой, что снижение микробного воздействия перестраивает иммунную систему таким образом, что на часто встречающиеся антигены окружающей среды быстрее развивается ответ Тh2-клеток. Однако эта гипотеза не подтверждена и механизмы наблюдаемого явления не установлены.
Таким образом, гиперчувствительность типа I представляет собой комплексное расстройство, возникающее в результате опосредованной IgE стимуляцией тучных клеток и последующим накоплением клеток воспаления в местах скопления антигена. Эти процессы регулируются главным образом индукцией Тh2-клеток, которые стимулируют продукцию IgE-антител (способствующих активации тучных клеток), вызывают скопление клеток воспаления (особенно эозинофилов) и секрецию слизи. Клинические признаки наблюдаются в результате высвобождения медиаторов тучными клетками и воспаления с присутствием большого количества эозинофилов.
В следующих статьях рассмотрим некоторые важные примеры IgE-опосредованных заболеваний.

Видео схема реакции гиперчувствительности I типа (аллергической реакции первого типа)
- Рекомендуем ознакомиться со следующей статьей "Примеры гиперчувствительности I типа (немедленного)"
Оглавление темы "Реакции иммунного ответа":- Цитокины иммунной системы
- Механизм презентации и распознавания антигенов лимфоцитами
- Механизмы клеточного иммунитета
- Механизмы гуморального иммунитета
- Механизмы затухания иммунного ответа и иммунологической памяти
- Механизмы и типы реакций гиперчувствительности
- Механизмы гиперчувствительности немедленного типа (I типа)
- Примеры гиперчувствительности I типа (немедленного)
- Механизмы гиперчувствительности опосредованной антителами (II типа)
- Механизмы гиперчувствительности опосредованной иммунными комплексами (III типа)