
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Транспортная РНК или тРНК. Генные аллели и их взаимодействие.
Гены тРНК умеренно повторены в геноме, их число измеряется в сотнях и тысячах: 850 — у дрозофилы, 1S50 — у шпорцевой лягушки, 1300 — у человека. У прокариот промотор примыкает к 5'-концу структурной части гена, а у эукариот промотор расчленен и находится внутри гена, один блок которого находится на расстоянии 8-30 п.н. левее инициирующего кодона, а второй — правее на +51 п.н.. В промоторе гена тРНК имеются А- и В-боксы. Установлены их канонические последовательности. При уменьшении расстояния между боксами транскрипция снижается или полностью прекращается. Замена всего одного нуклеотида в В-боксе в положении 56 CG -> GC в Tii/CG-петле искажает правильную структуру тРНК.
У прокариот некоторые кластеры с генами тРНК образуют опероны с общим промотором и одним предшественником зрелой РНК. Если есть интрон, расположенный, как правило, рядом с антикодоном, то он вырезается, как и в пре-мРНК. В митохондриях млекопитающих гены тРНК перемежаются со структурными генами.
Таким образом, генетический материал эукариот имеет три уровня организации: геномный, хромосомный и генный, каждый из которых обладает своими структурными особенностями. Гены являются единицей наследственности, они - составные элементы хромосом и генома в целом. Однако в геноме эукариот превалирует некоди-рующая ДНК (у человека —97%), функции которой изучены пока недостаточно. Тем не менее, установлена ее роль в альтернативном сплайсинге. Кроме того, в некоторых интронахобнаружены промоторы. Важное значение имеют регуляторные участки, расположенные с 5'- и 3'-концов гена, которые также относятся к некодирующей ДНК. Изучается роль тринуклеотидных повторов как в некодирующей так и в кодируюшей ДНК, увеличение числа которых выше нормы приводит к тяжелым заболеваниям.
Роль различных уровней организации генетического материала в передаче генетической информации потомкам в норме и патологии (когда генные, хромосомные и геномные мутации приводят к искажению этой информации) рассматриваются в различных главах пособия.
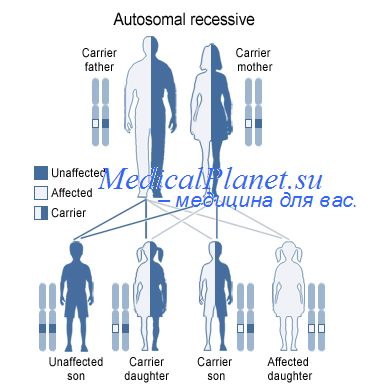
Генные аллели и их взаимодействие.
Аллель - одна из альтернативных форм гена. В классической генетике аллелями принято считать альтернативные фенотипические проявления признака у жизнеспособных особей. После открытия молекулярной структуры гена аллель стал трактоваться как один из вариантов нуклеотидной последовательности определенного фрагмента ДНК (как кодирующего, так и не кодирующего) в структуре генома. Например, аллелем называют и вариант длины рестрикционного фрагмента вне зависимости от его локализации, и последовательность варианта повтора сателлитной ДНК. Таким образом, в классической генетике к определению понятия аллель подходили с точки зрения его функционального проявления (различные варианты признака), а в молекулярной генетике учитывается прежде всего изменение структуры определенной последовательности ДНК.
При этом в случае некодирующей ДНК аллель, как правило, не имеет никакого фенотипического проявления. В этом, видимо, заключается защитная функция некодирующей ДНК от мутаций в ее структуре с фенотипическим проявлением. Вместе с тем показано, что изменения в последовательности оснований в регуляторных 5'- и 3'-нетранслируемых областях генов, нитронах и сайтах сплайсинга могут привести к тяжелым заболеваниям у человека.
Между аллелями одного гена существуют различные типы взаимодействия доминирование, неполное доминирование, кодоминирование, сверхдоминирование. В конце 70-х годов XX века Р. Ригером было предложено включить в эту классификацию еще два типа взаимодействия: неустойчивую доминантность и условную доминантность.
Термины «доминантный» и «рецессивный» признаки были введены Менделем в его работе «Опыты над растительными гибридами». Доминантные признаки могуг быть как нормальными, так и мутантными. Вне зависимости от локализации гена (в половой хромосоме или аутосоме) доминантные признаки проявляются в каждом поколении. Доминантный признак, контролируемый аутосомным геном, наследуется по аутосомно-доминантному типу, а при локализации гена в половой хромосоме - по сцепленному с полом доминантному типу. Различия этих двух типов наследования заключаются в том, что аутосомно-доминаптные признаки с одинаковой вероятностью передаются особям как мужского, так и женского пола, в то время как доминантные сцепленные с полом нормальные признаки или патологические мутации передаются от отца только дочерям. Если же родители оба больны и при этом мать гетерозиготна, то здоровыми могут быть только сыновья и то с вероятностью 50%.
Кроме того, доминантные мутации могут быть летальными в гомозиготном состоянии, они способны усиливать иди ослаблять действие других неаллельных генов, приводить к стерильности особей, одновременно влиять на различные признаки.
Как видно из этой таблицы доминантными могут быть не только морфологические признаки, но и биохимические, физиологические, поведенческие.
- Читать далее "Неполное доминирование в генетике. Кодоминирование в генных аллелях."
Оглавление темы "Взаимодействие генов.":1. Генетический материал. Генный уровень организации генетического материала.
2. Контроль матричных процессов генами. Рибосомная РНК.
3. Транспортная РНК или тРНК. Генные аллели и их взаимодействие.
4. Неполное доминирование в генетике. Кодоминирование в генных аллелях.
5. Неустойчивая и условная доминантность в генетике. Множественные аллели признака.
6. Механизмы доминантности и рецессивности генов. Контроль доминирования в аллелях.
7. Взаимодействие неаллельных генов. Комплементарность генов. Эпистаз генов.
8. Двойной рецессивный эпистаз. Характеристика и примеры рецессивного эпистаза.
9. Гены модификаторы. Взаимодействие эмбриональных генов.
10. Полигенное наследование признаков. Принципы полигенного наследования.