
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Контроль матричных процессов генами. Рибосомная РНК.
Биохимические процессы в клетке принято делить на матричные и нематричные. К матричным относят синтез ДНК, РНК и белков, их созревание и модификацию, а к нематричным - синтез низкомолекулярных соединений: аминокислот, азотистых оснований, Сахаров. В зависимости от того, какие именно процессы контролируют те или иные гены, их подразделяют на:
• гены, контролирующие структуру белков, которые участвуют в процессах синтеза аминокислот, азотистых оснований, Сахаров;
• гены, отвечающие за синтез рибосомной и транспортной РНК;
• гены, контролирующие синтез белков, которые обеспечивают матричные процессы: репликацию, транскрипцию и трансляцию.
По другой классификации гены делят на три класса в зависимости от типа РНК-полимеразы, которая их транскрибирует:
• гены рибосомных РНК: 18S, 5,8S, 28S, транскрибируемые РНК-полимеразой I;
• гены, кодирующие белки, транскрибируемые РНК-полимеразой Н;
• гены 5SpPHK, тРНК, мя (малых ядерных) РНК, транскрибируемые РНК-полимеразой III.
Гены рРНК организованы в мультигенное семейство. Число рибосомных генов 18S и 28S колеблется от 100 у мыши до 280 у человека и 780 у шпорцевой лягушки (Хепоpus). Для обеспечения синтеза многочисленных белков в клетке нужны рибосомы, а в их образовании участвуют различные фракции РНК. Рибосомные гены, как правило, локализованы в районе ядрышкового организатора (ЯОР). При нарушениях в области локуса данного гена у человека развивается ожирение.
После отхождения ядрышка от этого участка образуется вторичная перетяжка и спутник, который не всегда заметен при световой микроскопии. Кластеры генов: 18S-, 5,8S- и 28S-pPHK тандемно повторяются в районе ядрышкового организатора. Эти три гена разделены двумя транскрибируемыми спейсерами (ITS), имеются также внешние спейсеры (ETS) и межкластерные спейсеры (IGS) с промотором и терминатором.
У человека рибосомные гены 18S, 5.8S и 28S расположены в коротких плечах ак-роцентрических хромосом: 13,14,15,21,22;для них характерна также тандемная организация в районах ядрышковых организаторов. В геноме наблюдается индивидуальная и возрастная изменчивость по числу копий на хромосому.
Гены 5S-pPHK (120 п.н.) также умеренно повторены в геноме. У человека имеется около 2 000 таких генов. Причем они локализованы отдельно от других рибосомных генов. Фракция 5S-рРНК входит в состав большой субъединицы эукариотической рибосомы, гены этого типа, как правило, локализованы вне ядрышкового организатора.
Для генов 5S-рРНК характерны следующие особенности:
• как правило, они имеют кластерную организацию;
• в одну единицу повтора входит ген и его псевдоген
• гены 5S-рРНК в соматических клетках и ооцитах могут несколько отличаться по последовательности нуклеотидов;
• частота повтора генов 5S-pPHК достигает 2000 у человека и 24000 у лягушки Xenopus.
В состав эукариотического гена, кодирующего полипептидную цепь, входят: структурная часть и фланкирующие регуляторные участки с 5'и 3'-сторон.
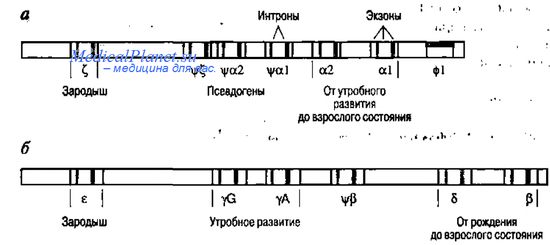
Структуру эукариотического гена рассмотрим на примере р-глобинового гена, состоящего из 3 экзонов и 2 интронов. Между первым и вторым экзоном расположен интрон IVS-1, а между вторым и третьим - IVS-2. Последовательности этих интронов идентичны у р-, у- и 5-генов, но отличаются от последовательности более коротких интронов у глобиновых генов.
В промоторе глобинового гена человека, наряду с TATA- и СААТ-боксами, расположенными на расстоянии от точки инициации транскрипции в 30 и 80 пар нуклеотидов, соответственно, имеется еще и третий элемент PuCPuCCC, где Pu - пурин. ACATTTG - кэп-последовательность кодирует 5'-конец РНК, к которому после синтеза первых трех десятков нуклеотидов присоединяется, модифицированный в 7-м положении метилированный гуанозин-5'-трифосфат. Между сайтами инициации транскрипции итрансляции расположена лидерная последовательность.
В глобиновом гене сайт терминации расположен через 1000 нуклеотидов от последнего экзона. Сигналом к разрезанию РНК-цепи служит появление участка AAUAAA. Сделав разрез примерно через 20 оснований после AAUAAA-мотива в молекуле РНК, поли-А-полимераза присоединяет «хвост» из 220 остатков адениловой кислоты. Однако транскрипция продолжается и после ААТААА-сайта до сайта терминации транскрипции, В этой 3'-концевой зоне (через 600-900 оснований после ААТААА-сайта) расположен энхансер, необходимый для организации экспрессии р-глобинового гена в предшественниках зрелых эритроцитов. С 5'-стороны за 6-22 т.п.н. до кластера Р-глобиновых генов расположена область, называемая LCR (от англ. locus activation region), включающая несколько эритроидспецифичных энхансеров, положение которых совпадаете положением участков гиперчувствительности к ДН Казе-1. В участках гиперчувствительности II- IV картированы многочисленные участки связывания различных транскрипционных факторов.
Гены глобинов человека образуют два мультигенных семейства. а-Глобиновые гены локализованы в коротком плече 16-й хромосомы в следующей последовательности: ген, кодирующий цепи эмбриональных гемоглобинов Gower 1 и Portland 1, затем три псевдогена - к 3'-концу в этом кластере-a2-l-гены. Кластер р-генов расположен в коротком плече 11-й хромосомы (11р) в следующей последовательности: е-ген эмбриональной цепи; два гена фетальных цепей гемоглобина HbF, которые отличаются по одному кодону; псев-доген цепи; 5-ген, кодирующий 5-цепь гемоглобина взрослого человека НbА2 и ген. В такой же последовательности эти гены начинают функционировать на различных этапах онтогенеза. Кластерная организация глобиновых генов напоминает оперонную структуру прокариот. Однако у прокариот в структуре оперона есть общие регуляторные последовательности: промотор, оператор, герминатор. В то время как каждый из глобиновых генов имеет свои регуляторные последовательности.
- Читать далее "Транспортная РНК или тРНК. Генные аллели и их взаимодействие."
Оглавление темы "Взаимодействие генов.":1. Генетический материал. Генный уровень организации генетического материала.
2. Контроль матричных процессов генами. Рибосомная РНК.
3. Транспортная РНК или тРНК. Генные аллели и их взаимодействие.
4. Неполное доминирование в генетике. Кодоминирование в генных аллелях.
5. Неустойчивая и условная доминантность в генетике. Множественные аллели признака.
6. Механизмы доминантности и рецессивности генов. Контроль доминирования в аллелях.
7. Взаимодействие неаллельных генов. Комплементарность генов. Эпистаз генов.
8. Двойной рецессивный эпистаз. Характеристика и примеры рецессивного эпистаза.
9. Гены модификаторы. Взаимодействие эмбриональных генов.
10. Полигенное наследование признаков. Принципы полигенного наследования.