
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Мейоз. Фазы мейоза. Процессы во время мейоза.
Мейоз форма деления характерна для специализированных клеток половых желез (яичников и семенников). Издиплоидных клеток-предшественников в ходе мейоза образуются гаплоидные гаметы — яйцеклетки и сперматозоиды (число хромосом п). Мейоз включает два клеточных деления, перед которыми происходит только одна репликация ДНК. Первое деление называется редукционным и обозначается как мейоз I. В результате этого деления из одной диплоидной клетки образуются две гаплоидные. Второе деление называется эквационным и обозначается как мейоз II. Это деление подобно митозу, так как сестринские хроматиды отделяются друг от друга и расходятся к разным полюсам. Каждое из двух делений мейоза состоит из профазы, метафазы, анафазы и телофазы.
МЕЙОЗ 1. Во время профазы I происходитспирализация и укорочение хромосом. Кроме того, гомологичные хромосомы конъюгируют друг с другом по всей длине, образуя бивалент Во время конъюгации хромосом между несестринскими хроматидами может произойти обмен участками - кроссинговер. В точке обмена образуется видимая в световой микроскоп крестообразная структура, которую называют хиазм ой. Генетические исследования свидетельствуют о том, что кроссинговер происходит на стадии четырех хроматид, при этом в данной точке обмениваются участками только две из четырех нитей, и хроматиды участвуют в обмене случайно. Как правило, чем больше длина хромосом, тем больше среднее число образуемых ими хиазм. В электронном микроскопе хорошо видно, как между гомологичными хромосомами в каждом биваленте формируется особая структура — синаптонемный комплекс, но в некоторых случаях этот комплекс не образуется и в результате кроссинговер или отсутствует, или сильно подавлен. В частности, синаптонемный комплекс не выявляется у самцов дрозофилы, не происходит у них и кроссинговер. Описаны мутации, приводящие к отсутствию синаптонемного комплекса в профазе I у самок дрозофилы. Например, у самок, гомозиготных по мутации c(3)G, полностью подавлен кроссинговер. Электронномикроскопические исследования показали, что у них не формируется синагпонемный комплекс.
По морфологии ядра и хромосом, наблюдаемой в световой микроскоп, профазу первого деления мейоза можно разделить на ряд стадий:
1) лептотену (стадия тонких нитей) - в ядре начинают выявляться длинные тонкие нити хромосом вместо гранул хроматина интерфазного ядра;
2) зиготену (стадия объединения нитей) - начинается конъюгация гомологичных хромосом;
3) пахитену (стадия толстых нитей) - синапс хромосом настолько тесный, что отдельные гомологи в биваленте неразличимы;
4) диплотену (стадия двойных нитей) — синапс становиться менее тесным, уже видны хроматиды и хиазмы;
5) диакинез — хромосомы максимально укорачиваются, центромеры гомологичных хромосом отталкиваются друг от друга, и хромосомы удерживаются вместе только в зонах хиазм, исчезают ядрышко и ядерная мембрана, начинает формироваться веретено деления.
Метафаза I. Биваленты выстраиваются в экваториальной плоскости, при этом центромеры ориентируются относительно полюсов случайно.
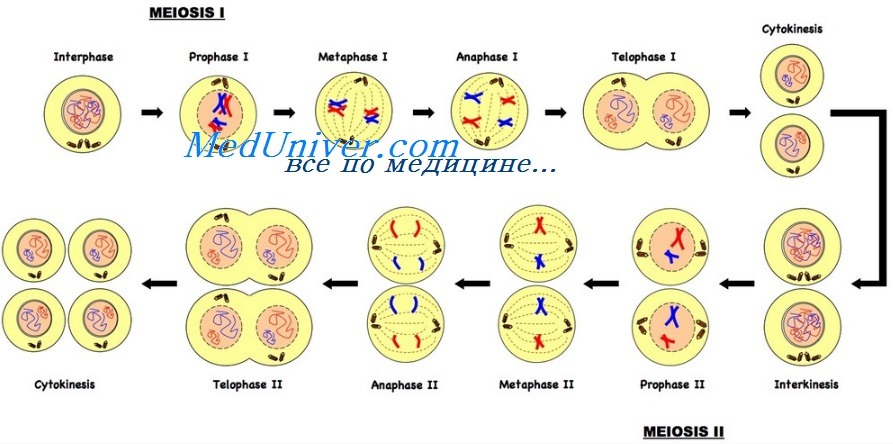
Анафаза I. Гомологичные хромосомы отделяются друг от друга и движутся к противоположным полюсам. Центромеры не расщепляются, поэтому сестринские хроматиды продолжают удерживаться вместе. Вследствие кроссинговера сестринские хроматиды уже могут быть неидентичными. Таким образом, к полюсам движутся хромосомы, состоящие из двух хроматид, В ходе этой фазы из одной диплоидной клетки образуется две гаплоидные.
Интеркинез. Это стадия между первым и вторым делениями мейоза. У разных видов интеркинез имеет разную продолжительность. Если эта стадия длительна, хромосомы могут деком пакта зоваться и принять вид интерфазного хроматина. Важно помнить, что на этой стадии не происходит репликации ДНК.
МЕЙОЗ II. В профазе И восстанавливается веретено деления. Во время метафазы II хромосомы располагаются в экваториальной плоскости. В анафазе II происходит расщепление центромер, и хроматиды каждой хромосомы разделяются и движутся к противоположным полюсам деления. Втелофазе II из каждого гаплоидного ядра образуются два, которые содержат гаплоидное число хромосом, состоящих из одной хроматиды каждая.
Таким образом, диплоидная родительская клетка делится на четыре гаплоидные. Генетаческое содержимое этих клеток различно. Материнские и отцовские хромосомы могут находиться в них в разных комбинациях, при этом в результате кроссинговера в каждой хромосоме также могут появиться новые комбинации аллелей.
При слиянии разнообразных гамет в процессе оплодотворения возникают новые диплоидные потомки с различными комбинациями отцовских и материнских генов.
В 1902 г. У. Сэттон и Т. Бовери, сопоставляя поведение хромосом в мейозе и поведение признаков при передаче их от родителей потомкам, пришли к предположению, что менделевские наследственные факторы (гены) расположены в хромосомах. Их гипотеза основывалась на сходстве поведения генов и хромосом при образовании гамет и зигот.
1. В ядре каждой соматической клетки содержится по две гомологичные хромосомы. Каждая соматическая клетка несет по два аллеля одного гена,
2. Гомологичные хромосомы в профазе мейоза конъюгируют и в норме обязательно расходятся к противоположным полюсам веретена деления. Следовательно, гаметы несут только по одной хромосоме из каждой пары. Также и аллели одного гена при образовании гамет обязательно попадают в разные гаметы, каждая гамета несет только один из аллелей (правило чистоты гамет).
3. При оплодотворении из гаплоидных гамет образуется диплоидная зигота, восстанавливается диплоидное число хромосом. Одну из гомологичных хромосом зигота получает от матери, вторую — от отца. То же известно и о генах: каждый организм имеет два аллеля одного гена, причем один аллель получен от матери, второй - от отца.
4. Ориентация негомологичных хромосом к полюсам случайна, гаметы могут иметь все возможные комбинации хромосом: как все отцовские или все материнские, так и разные сочетания отцовских и материнских хромосом. То же известно и о генах: неаллельные гены наследуются независимо.
Параллелизм поведения хромосом и генов в процессе клеточного деления указывает на то, что гены — составная часть хромосом.
Доказательства хромосомной локализации генов были получены Т Морганом (1910 г.) и К. Бриджесом (1916 г.) при изучении наследования признаков, сцепленных с полом, у дрозофилы.
Учебное видео: мейоз и его фазы
- Читать далее "Половые различия в типе хромосом. Признаки сцепленные с полом."
Оглавление темы "Хромосомы и гены.":1. Хромосомы. Хромосомная теория наследственности.
2. Клеточный цикл. Хромосомы в ходе клеточного цикла. Митоз и фазы митоза.
3. Мейоз. Фазы мейоза. Процессы во время мейоза.
4. Половые различия в типе хромосом. Признаки сцепленные с полом.
5. Нерасхождение половых хромосом. Изучение нерасхождения половых хромосом.
6. Группы сцепления генов в хромосоме. Понятие группы сцепления.
7. Теория наследственности Моргана. Аутосомно-доминантное наследование признаков.
8. Аутосомно-рецессивный тип наследования признаков. Недоминантное наследование.
9. Ген. История изучения генов. Роль гена в мутации и наследственности.
10. Молекула ДНК. Открытие ДНК. Генетическая роль ДНК.