
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Цитоскелет и клеточная динамика клетки - с точки зрения физиологии человека
Оглавление статьи:- Каркас живой клетки
- Отравление колхицином
- Моторные белки
- Перемещение клеток
- Список использованной литературы
а) Каркас живой клетки. Всем клеткам в процессе их роста, деления, адаптирования к новой среде постоянно нужна реорганизация их содержимого; эту задачу выполняет цитоскелет — динамическая система филаментов.
Цитоскелет включает в себя три основных компонента: актиновые филаменты, микротрубочки и промежуточные филаменты (рис. 1).
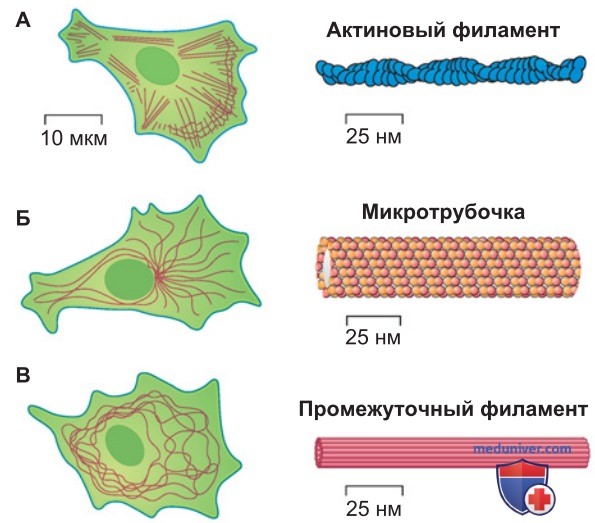
1. Актин. Актиновые филаменты представляют собой двухцепочечные спиралевидные полимеры белка актина. Это гибкие структуры диаметром 4-9 нм в виде линейных пучков, двухмерных сетей и трехмерных гелей.
Хотя актиновые филаменты встречаются в клетке повсеместно, сосредоточены они непосредственно под клеточной мембраной, кортексом клетки. Актиновые филаменты определяют форму клетки и играют решающую роль в движении клеток. Однако во взаимодействии актина с другими конструктивными элементами клетки участвует много дополнительных (вспомогательных) белков, в том числе моторные белки, которые перемещают либо органеллы вдоль филаментов, либо сами филаменты.
2. Микротрубочки. Это длинные неветвящиеся цилиндры из белка тубулина. Их наружный диаметр ~25 нм, и они намного прочнее, чем актиновые филаменты. Все микротрубочки одним своим концом прикреплены к центросоме (клеточному «центру организации микротрубочек»), от которой они берут начало. Обычно центросома расположена вблизи ядра.
Микротрубочкам принадлежит исключительная роль в клеточном делении: они образуют биполярное митотическое веретено, в средней части которого находятся хромосомы. Кроме того, они могут формировать подвижные клеточные выросты (реснички) на поверхности клеток, а также создают вдоль аксонов длинные прямые стержни, которые обеспечивают перемещение материала из тела клетки (сомы) на периферию (аксонный транспорт).
3. Промежуточные филаменты. Это волокна диаметром около 10 нм, перевитые наподобие веревки. Они принадлежат большому разнородному семейству и построены из различных белковых молекул. На внутренней стороне оболочки ядра клетки промежуточные филаменты образуют густую сеть, так называемую ядерную пластинку, которая охватывает ДНК наподобие защитной оболочки.
Кроме того, из промежуточных филаментов состоит крупноячеистая цитоплазматическая сеть, придающая клетке механическую прочность. В эпителии такая сеть распространяется даже между клетками, обеспечивая эпителиальной ткани очень высокую надежность. Примерами могут служить ситуации, требующие значительного растяжения ткани: прохождение пищи через кишечник (задействованы клетки слизистой оболочки кишечника), опорожнение мочевого пузыря (эпителиальные клетки мочевого пузыря), увеличение площади кожи живота во время беременности (клетки эпидермиса).
б) Отравление колхицином. Колхицин — яд безвременника осеннего (Colchicum autumnale). Отравление случается большей частью у детей (которые берут в рот стебли растения) либо в результате терапевтической передозировки при лечении подагры.
1. Патология. Связываясь с белками микротрубочек, колхицин ингибирует внутриклеточный транспорт и деление клеток (митотический яд).
Терапевтическое применение при подагре может сопровождаться накоплением мочевой кислоты в организме. В низких дозах колхицин подавляет захват фагоцитами кристаллов мочевой кислоты в тканях, подавляя таким образом воспалительный процесс.
2. Побочное действие. В более высоких дозах колхицин обладает антимитотическим действием, которое прежде всего затрагивает быстро делящиеся клетки эпителия и кроветворной системы. Это приводит к кровотечениям, диарее и нарушениям дыхания.
в) Моторные белки. Моторные белки ассоциированы с цитоскелетом и транспортируют необходимые материалы в определенные участки клетки; транспорт происходит с затратой энергии, которую поставляет АТФ.
1. Молекулярные двигатели. Моторные белки — замечательные в своем роде молекулы, связанные с цитоскелетом (рис. 2). Снабжаемые энергией АТФ, они передвигаются вдоль филаментов цитоскелета. Существуют десятки различных моторных (двигательных) белков. Они различаются тем, что связываются с филаментом лишь одного типа, движутся по клетке только в конкретных направлениях, транспортируют определенный материал.
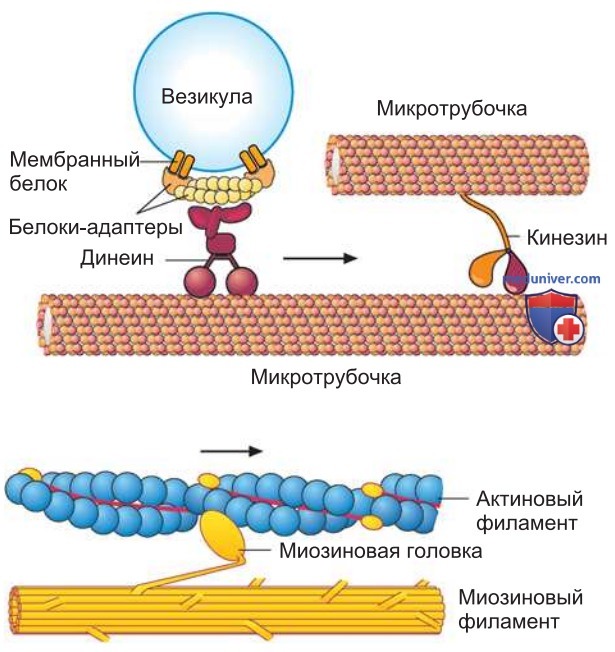
Одни моторные белки перемещают митохондрии, стопки мембран аппарата Гольджи и секреторные везикулы на свойственные им места. Другие изменяют относительное расположение филаментов цитоскелета; в результате развивается механическое усилие, которое в конечном счете приводит к сокращению мышц, биению ресничек или делению клеток.
Моторные белки цитоскелета присоединяются к соответствующим филаментам посредством головного домена, который связывается с АТФ, инициируя его гидролиз. Гидролиз АТФ сопровождается конформационными изменениями моторных белков. В зависимости от конформации белок связывается с филаментом или же освобождается от него. Таким способом моторный белок постепенно «едет» вдоль филамента. Головной домен молекулы задает направление, хвостовой регион — вид транспортируемого материала.
2. Миозин. Это первый моторный белок, получивший известность. Он генерирует силу, необходимую для мышечного сокращения. При связывании длинной «двухголовой» молекулы этого белка происходит гидролиз АТФ и филамент миозина скользит вдоль актинового филамента. Нужно отметить, что миозин обнаружен не только в мышечных клетках. Это целое молекулярное семейство, насчитывающее более дюжины представителей. Многообразные функции отдельных типов миозина до сих пор не вполне выяснены.
3. Кинезин. Моторный белок, который двигается вдоль микротрубочек. Его молекула несет две головки и сходна по своей структуре с мышечным миозином (миозином II). Кинезин принадлежит к обширному суперсемейству белков. Хвостовой регион большинства кинезинов имеет участок связывания для окруженных мембранами органелл либо для других микротрубочек. Многие члены этого суперсемейства выполняют важную роль в формировании митотического веретена и в расхождении хромосом при делении клетки.
4. Динеины. Это самые крупные из известных к настоящему времени моторных белков. В семействе динеинов можно выделить две основные группы. Цитоплазматические динеины транспортируют везикулы по клетке и закрепляют аппарат Гольджи в клеточном центре. Другие динеины осуществляют быстрое скольжение микротрубочек при биении ресничек, например в эпителии дыхательных путей. Динеины относятся к самым быстрым молекулярным моторам. Они способны передвигать микротрубочки со скоростью 14 мкм/с, тогда как кинезины обеспечивают скорость максимум 2-3 мкм/с.
г) Перемещение клеток. Клетки перемещаются в нашем организме иногда на значительные расстояния; форма движения зависит от типа клетки.
1. Ползание клеток. Миграция — важный физиологический и патофизиологический процесс в жизни клетки (рис. 3). Уже в ранний период эмбриогенеза клетки переползают на большие расстояния. В результате миграции клеток нервной трубки формируется нервная система эмбриона. Зародышевые нервные клетки (нейробласты), родившиеся в центральной нервной системе, перемещаются к своим окончательным рабочим позициям. Белые кровяные клетки (лейкоциты) «охотятся» за проникшими в организм бактериями и другими патогенами либо перемещаются в очаг воспаления.
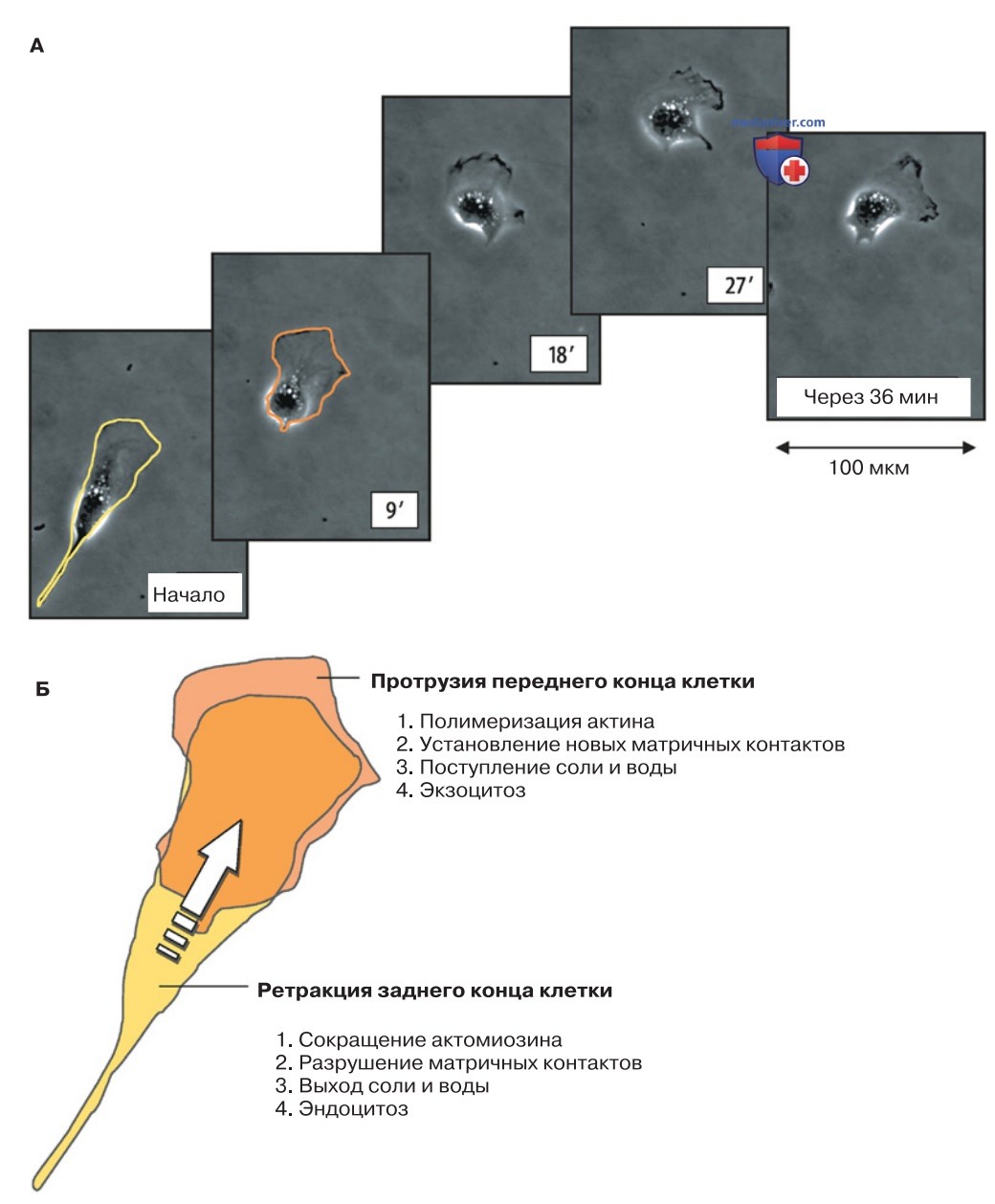
Клетки соединительной ткани (фибробласты) выходят в раны, обеспечивая их заживление (образование рубцов). Аналогичным образом ведут себя эпителиальные клетки, которые занимают места предшествующих клеток после их отмирания. Для роста и развития кровеносных сосудов (ангиогенеза) необходима миграция клеток эндотелия. И наконец, если мы обратимся к патофизиологическим аспектам, то увидим, что в результате подвижности раковых клеток образуются метастазы (вторичные новообразования) и таким образом опухоль распространяется в организме.
2. Скорость перемещения клеток значительно варьируется в зависимости от их типа. Эпителиальные клетки мигрируют со скоростью 0,10,2 мкм/мин, лейкоциты — до 5-10 мкм/мин, а некоторые клетки кожи достигают скорости до 30 мкм/мин. Несмотря на эти различия, механизмы миграции сходны для всех клеток нашего организма. В типичном случае на поверхности клетки формируются два полюса движения. «Передний» полюс клетки — ламеллоподия, пластинка толщиной ~300 нм, лишенная органелл. Противоположный «задний» полюс образован телом и хвостовой частью клетки.
3. Полимеризация актина. Передвижение клетки по твердому субстрату — сложный процесс, главную роль в котором играет гелеобразный кортикальный слой актиновых филаментов (кортекс), лежащий непосредственно под клеточной мембраной. Первый этап поступательного движения клетки соответствует полимеризации актина. При этом актиновые филаменты упираются в плазматическую мембрану и выпячивают ее наружу, создавая подвижные выступы клеточной поверхности — филоподии (микрошипы) и ламеллоподии (тонкие пластинчатые отростки), необходимые для передвижения клетки (локомоции).
Если в эксперименте эти выросты отделить от клетки, они могут продолжать движение самостоятельно. Таким образом в результате циклов полимеризации—деполимеризации актина, при участии других моторных белков и с затратой энергии гидролиза АТФ в конечном счете может происходить направленное движение.
4. Поступательное движение клеток. Это движение обеспечивают по меньшей мере четыре молекулярных процесса.
- При временном локальном поступлении Ca2+, других ионов и воды происходит разжижение богатого актином геля под клеточной мембраной, так что ламеллоподия обретает подвижность. Когда концентрация Ca2+ снижается и вода выходит из клетки, актин полимеризуется и толкает ламеллоподию вперед.
- Одновременно на противоположном конце клетки плазматическая мембрана инвагинирует и от нее отпочковываются внутриклеточные эндоцитозные везикулы. Они транспортируются вдоль микротрубочек к передней части клетки, или «ведущему краю» (leading edge), где встраиваются в липидный бислой плазматической мембраны (lipid flow).
- В передней части клетки посредством специфических транспортных механизмов (Na+/H+-антипорта, Cl-/HCO-3-антипорта) поглощается NaCl вместе с водой. «Ведущий край» увеличивается в объеме и продвигается вперед.
- На противоположном, заднем, конце клетки ионы выходят из нее вместе с водой через каналы. При этом конец клетки укорачивается.
Клетка выстраивает перед собой участок внеклеточного матрикса (матричные белки), по которому продвигается вперед, как по асфальтированной улице. Получая энергию за счет процессов, упоминавшихся выше, клетка прикрепляется к внеклеточному матриксу, связываясь с белками интегринами. Такое связывание — фокальные контакты (focal contacts) — носит локальный и кратковременный характер.
Благодаря координированным молекулярным процессам осуществляется скользящее движение. Его направление определяют сигнальные вещества внешней среды (хемотаксис). Так, лейкоцит движется непосредственно к бактерии, поскольку она выделяет специфичные белковые молекулы-аттракторы.
д) Коротко. Цитоскелет и клеточная динамика. Форма и деятельность клетки зависят от динамических элементов ее структуры. Актиновые филаменты в основном располагаются непосредственно под плазматической мембраной. Они то образуются, то снова распадаются на отдельные молекулы, определяя подвижность всей клетки. Это служит предпосылкой для физиологически значимого передвижения клеток (миграции).
Значительно более толстые микротрубочки представляют собой полые цилиндры, которые при клеточном делении упорядочивают положение хромосом и способствуют перемещению внутриклеточных компонентов. Механическую стабильность клеточного ядра обеспечивает лежащая под ядерной оболочкой густая сеть прочных промежуточных филаментов. За пределами ядра эти филаменты тянутся через всю клетку, поддерживая ее структуру в случае растяжения. К поверхности филаментов прикреплены моторные белки (миозин, динеин и кинезин); они опосредуют доставку определенных материалов к соответствующим участкам клетки.
Видео строение и функции цитоскелета
- Рекомендуем ознакомиться далее "Процессы поглощения и выделения клетки - с точки зрения физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 1.7.2024