
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Циклические нуклеотиды в роли вторичных мессенджеров клетки - с точки зрения физиологии человека
Оглавление статьи:а) Циклический аденозинмонофосфат. С помощью аденилатциклазы образуется циклический аденозинмонофосфат (цАМФ), который активирует протеинкиназу А и может оказывать влияние на эффекторные молекулы и процесс экспрессии генов; в результате этого цАМФ снова инактивируется под воздействием фосфодиэстеразы.
1. Аденилатциклаза. Активированные α-субъединицы определенных гетеротримерных G-белков (Gs) взаимодействуют с аденилатциклазой, которая преобразует АТФ в циклический АМФ (рис. ниже). цАМФ — внутриклеточный медиатор (вторичный мессенджер), который осуществляет действие гормона (первичный посредник) в клетке.
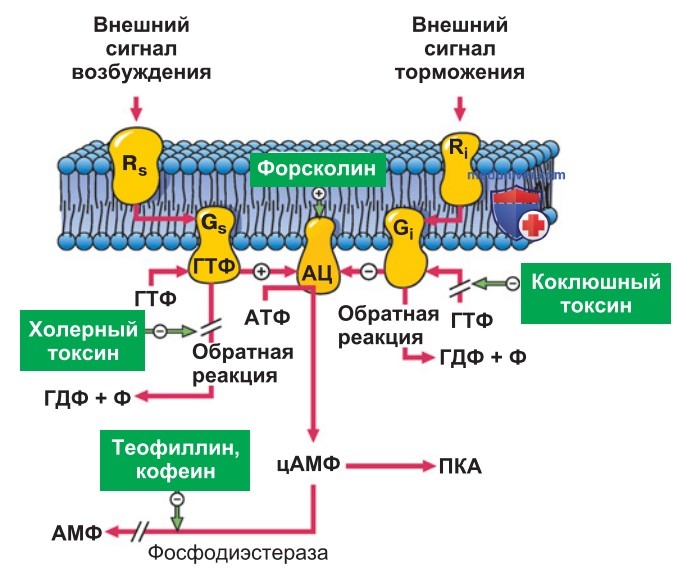
цАМФ присоединяется к протеинкиназе А (ПКА) и активирует ее. ПКА фосфорилирует определенные ферменты, ионные каналы и другие транспортные белки в серине или треонине, оказывая тем самым влияние на их функции. цАМФ может накапливаться в ионных каналах и активировать их без участия ПКА.
ПКА фосфорилирует фактор транскрипции белок CREB (cAMP responsive element binding protein) и вызывает экспрессию цАМФ-зависимых генов. В результате цАМФ может активировать ионные каналы напрямую.
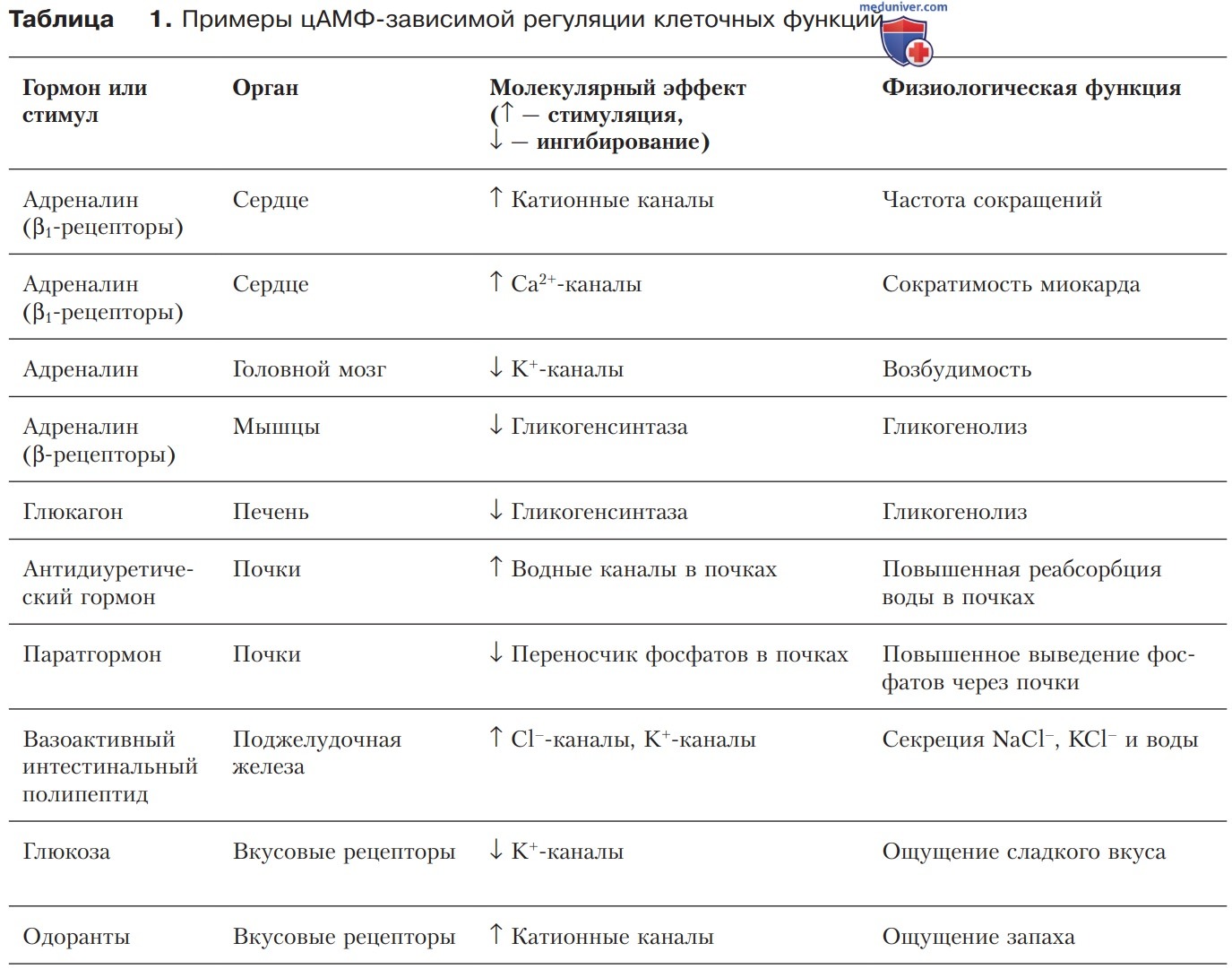
Большое количество гормонов, например адреналин (через β-рецепторы), глюкагон, паратгормон, кальцитонин, большинство пептидных гормонов таламуса и гипоталамуса (за исключением соматостатина) и другие тканевые гормоны, передают сигнал описанным здесь образом. Некоторые примеры цАМФ-зависимой регуляции представлены в табл. 1.
2. Инактивация. Под воздействием фосфодиэстеразы происходят расщепление цАМФ до 5'-АМФ и его инактивация. Ингибирование фосфодиэстеразы (в частности, кофеином) приводит к повышению концентрации цАМФ в цитозоле и тем самым способствует выполнению функций клетки, зависимых от цАМФ (кофеин оказывает влияние преимущественно при стимуляции пуринергических рецепторов).
Под воздействием серин/треонинфос-фопротеинфосфатазы (РР1, РР2a, b, c) фосфорилирование белков останавливается, в результате чего действие ПКА блокируется.
3. Холерный токсин. Возбудитель холеры — холерный вибрион — продуцирует холерный токсин, вызывающий перемещение группы АДФ-рибозы в Gsα-субъединицу G-белка. При этом ГТФаза ингибируется, но G-белок остается в активном состоянии. В результате возникает очень сильная и продолжительная активация аденилатциклазы в эпителии кишечника.
Повышенное образование цАМФ приводит к открыванию хлорных каналов в люминальной мембране эпителия кишечника. Из-за усиленного выделения NaCl и воды возникает диарея, которая приводит к угрожающей жизни потере жидкости.
4. Торможение образования цАМФ. Гетеротримерные G-белки могут не только активировать ПКА, но и тормозить ее. Для этого рецептор взаимодействует с одним из ингибированных Gi-белков. После отщепления ГТФ и распада соединения субъединиц α, β и γ G-белки тормозят выделение аденилатцикла-зы, а клеточная концентрация цАМФ и активность ПКА соответственно снижаются.
Эти механизмы используются такими гормонами, как ацетилхолин, соматостатин, ангиотензин II или адреналин (через α2-рецепторы). Затормаживая образование цАМФ, соматостатин может препятствовать выделению Cl-, а адреналин — выбросу инсулина.
б) Циклический гуанозинмонофосфат. Гуанилатциклаза образует циклический гуанозинмонофосфат (цГМФ), который оказывает влияние на функции клетки при участии G-киназы; цГМФ помогает функционировать оксиду азота (NO) — чрезвычайно быстро распадающемуся медиатору.
1. Рецепторы гуанилатциклазы. Лишь некоторые рецепторы соединяются с гуанилатциклазой, которая высвобождает цГМФ из ГТФ, а цГМФ присоединяется к протеинкиназе G, оказывающей влияние посредством фосфорилирования белка. Среди прочих она активирует Са2+-АТФазу, которая выкачивает Са2+ из клетки; при участии цГМФ на нее воздействует в том числе и атриопептин. цГМФ также может присоединяться к ионным каналам и тем самым регулировать их активность.
Активируемый с помощью цГМФ катионный канал регулирует активность, например, зрительных рецепторов.
2. Оксид азота. Так называемые растворимые гуанилатциклазы регулируются не через рецепторы, а под воздействием оксида азота (NO), который образуется в клетке из аргинина при участии NO-синтаз (NOS). NOS в эндотелиальных клетках (eNOS) и мозге (nNOS) активируются с помощью Са2+. При воспалении выделяется индуцируемая NOS (iNOS), не требующая повышенной концентрации Са2+ в цитозоле для активации.
Будучи очень нестабильным соединением, NO предназначен для осуществления краткосрочных эффектов и способен модифицировать функции белков посредством нитрозации. Прежде всего он имеет большое значение при регуляции тонуса сосудов и передаче сигналов нейронам, однако играет важную роль и при регуляции генетически запрограммированной гибели клетки.
в) Коротко. Циклические нуклеотиды. Многие гормональные рецепторы регулируют деятельность клеток посредством циклических нуклеотидов, выполняющих функцию вторичных мессенджеров. Циклический аденозинмонофосфат (цАМФ) активирует протеинкиназу А и тем самым получает возможность оказывать влияние на молекулярные эффекторы и экспрессию генов. Циклический ГМФ (цГМФ) оказывает действие на функции клетки с помощью G-киназы. Концентрацию обоих вторичных посредников цАМФ и цГМФ регулирует активность аденилатциклазы или гуанилатциклазы.
- Вернуться в раздел "Физиологии человека"
Редактор: Искандер Милевски. Дата публикации: 1.7.2024