
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Контроль синтеза коллагена и внеклетомного матрикса (ВКМ)
Основные биологические особенности коллагена связаны с механизмом, контролирующим накопление коллагена в тканях. Такой контроль должен существовать in vivo, поскольку последствия бесконтрольной аккумуляции коллагена проявляются заболеваниями, в частности прогрессирующим системным склерозом и различными другими нарушениями фиброзного характера.
Накопление коллагена в тканях контролируется на нескольких различных уровнях биосинтеза и распада. Некоторые наблюдения указывают на то, что важный контрольный механизм действует на этапе образования матричной РНК (мРНК) и осуществляется путем регулировки транскрипционной активности генной экспрессии. В транскрипционной регулировке экспрессии генов участвуют как cis-действующие элементы, так и транс-действующие факторы.
Cis-действующие элементы представляют собой нуклеотидные последовательности в промоутерном участке гена, которые служат местами связывания транс-действующих клеточных белков, стимулирующих или подавляющих транскрипционную промоутерную активность. Некоторые из транс-действующих факторов являются ядерными рецепторами, например, рецепторы ретиноевой кислоты (RAR и RXR), которые образуют комплекс с лигандом (ретиноидом) и затем связываются с реагирующими на ретиноевую кислоту элементами (RARE) в гене-мишени.
Ретиноиды, такие как полностью транс-ретиноевая кислота, модулируют экспрессию гена коллагена типа I как in vivo, так и in vitro. В частности, покоящиеся, не пролиферирующие клетки могут стимулироваться ретиноевой кислотой и активировать экспрессию гена коллагена. Это может иметь отношение к повышенному синтезу коллагена в фотоповрежденной коже после местного применения полностью-транс-ретиноевой кислоты.
Различные факторы роста, цитокины и хемокины высвобождаются во внеклеточное пространство из разных типов клеток, таких как фибробласты, кератиноциты, эндотелиальные и воспалительные клетки. Путем паракринных и аутокринных механизмов эти факторы воздействуют на внутриклеточные сигнальные пути, действующие на транкрипционном и/или трансляционном уровнях.
Одним из наиболее мощных регуляторов производства коллагенов и внеклеточного матрикса является трансформирующий фактор роста β (TGF-β), член семейства факторов роста, участвующих во многих физиологических и патологических состояниях. Интересно отметить, что TGF-β может инициировать экспрессию других факторов роста, в частности фактора роста соединительной ткани в фибробластах.
Таким образом, считается, что производство коллагенов индуцируется цитокинами, такими как фактор роста тромбоцитов, интерлейкины-1 и -4 и хемокинами, такими как моноцитарные хемотаксические белки 1 и 3.
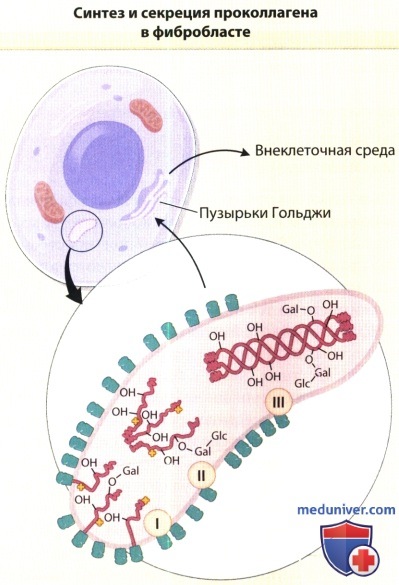
В увеличенном изображении представлены процессы, происходящие в шероховатой эндоплазматической сети клеток в ходе синтеза проколлагена.
На первом этапе (I) полипептидные цепи проколлагена синтезируются на мембранносвязанных рибосомах, и формирующиеся цепи направляются в цистерны шероховатой эндоплазматической сети.
На растущих полипептидных цепях начинается гидроксилирование пролиловых и лизиловых остатков и гликозилирование гидроксилизиловых остатков.
Эти реакции завершаются вскоре после высвобождения полноразмерных цепей из рибосом (II).
В процессе образования межцепочечных дисульфидных связей три про-а-цепи соединяются, и коллагеновые участки полипептидов приобретают конфигурацию тройной спирали (III).
Затем молекулы проколлагена перемещаются из шероховатой эндоплазматической цепи в пузырьки аппарата Гольджи и секретируются из пузырьков во внеклеточную среду.
— ОН = гидроксильные группы гидроксипролина и гидроксилизина; -Gal = галактозиловый остаток, присоединенный к гидроксилизину;
-Gal-Glc = глюкозилгалактозиловый остаток, присоединенный к гидроксилизину О-гликозидной связью; структуры в форме листа клевера обозначают гидроксилирующие и гликозилирующие ферменты.
- Вернуться в оглавление раздела "дерматология."
Оглавление темы "Внеклеточный матрикс (ВКМ) дермы.":