
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Эукариотический геном. Повторы или избыточность генома.
Геном эукариот значительно больше по своим размерам чем геном прокариот. Так максимальный размер генома прокариот не превышает 8*106 п. и., в то время как у дрожжей он равен 1,35*107, у шелкопряда 5*108, у шпорцевой лягушки, мыши и человека - 3*109, У некоторых растений и амфибий геном достигает 1011 п.н. Из приведенных примеров видно, что размер генома не прямо зависит от систематического положения организма. Считается, что геномы высших организмов содержат большой избыток ДНК, Так геном млекопитающих содержит не менее 3*109 п.н., что достаточно для кодирования 3 млн. белков средних размеров. Однако по расчетам, количество белков у млекопитающих составляет несколько десятков тысяч. Геном человека насчитывает около 3,3 млрд. нуклеотидных пар (гаплоидный набор), что соответствует примерно 3,5 пикограммам ДНК, при этом на долю кодирующих областей приходится не более 3%.
Повторы генома. Последовательности, присутствующие в геноме в виде нескольких копий называют повторяющимися, или повторами, а количество копий в геноме — частотой повторяемости. Наличие повторов - отличительная особенность эукариотических геномов, которая выявляется в экспериментах поденатурапии-ренатурацииДНК. Если ДНКденатурировать (разделить на фрагменты) а затем создать условия для ренатурации, или реассоциации, то скорость реассоциаиии будет зависеть от количества копий, или концентрации соответствующих фрагментов. Чем больше в геноме высокоповторяющихся последовательностей, тем менее сложно устроена его структура и тем быстрее идет ренатурация. В отличие от прокариот, геном которых представлен практически только уникальными последовательностями ДНК, в составе эукариот обнаруживаются фракции ДНК, которые реассоциируют значительно быстрее остальных (высокоповторяюшиеся участки). Кроме того, имеются и среднеповторяющиеся фрагменты с более медленной кинетикой реассоциации, но более высокой, чем у уникальных последовательностей ДНК. Более детальный анализ позволяет выявить и слабоповторяющиеся фрагменты (последовательности, представленные в виде нескольких десятков копий).
У человека высокоповторяющаяся ДНК составляет примерно 15% всей ДНК, среднеповторяюшаяся — 8% генома, слабоповторяюшиеся фрагменты (десятки и сотни копий) - около 4,4% генетического материала человека (эти цифры не совпадают у разных авторов и по мере дальнейших исследований могут быть изменены).
Все повторы можно подразделить на тандемно повторяющиеся, т.е. расположенные друг за другом «голова к хвосту», и диспергированные по геному. Тандемные повторы встречаются между генами и в нитронах, но больше всего их в центромерных и теломерных районах хромосом. При денатурации суммарной геномной ДНК и последующей реассоциации длинные тандемные повторы составляют фракцию быстро ренатурирующей ДНК. При равновесном центрифугировании в градиенте плотности CsCl эти повторяющиеся элементы образуют зоны разной плотности. Минорный компонент ДНК, отделяющийся от основной ДНК при равновесном центрифугировании в градиенте плотности хлористого цезия, принято называть сателлитной ДНК. Последовательности повторяющихся единиц сателлитной ДНК (сатДНК) видоспецифичны. Так у Drosophila melanogaster имеются четыре основных типа сателлитных последовательностей, состоящие из 5, 7, 10 и большего числа нуклеотидов, вместе составляющих 16% геномной ДНК. ДНК Drosophila virilis имеет три основных сателлита, каждый из которых состоит из 7 п.н., незначительно различающихся между собой по составу азотистых оснований.
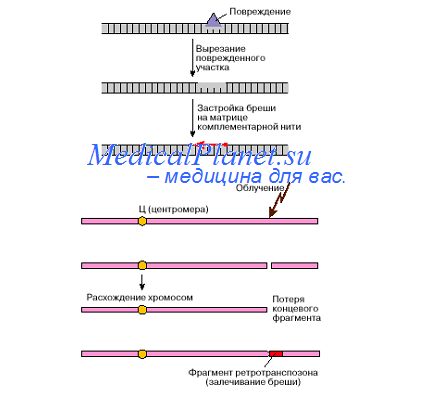
У млекопитающих, например, у грызунов, небольшую часть копий составляют идентичные последовательности, остальные отличаются множеством замен оснований, делений и вставок. Помимо видоспецифичности и отличий повторов сателлитной ДНК по составу оснований, для некоторых вариантов тандсмньгх повторов характерна специфичность их локализации в хромосомах. Так, декамсрные повторы второго сателлита Drosophila melanogaster локализованы в X и Y-хромосомах, а первый сателлит встречается в X, Y и 4-й хромосомах. У дрозофилы фракция, осаждающаяся взоне 1,679 г/см3, включает гены рРНК. Однако гены рибосомной РНК, умеренно повторены в геноме и транскрибируются с образованием различных фракций рРНК, в то время как сателлитная ДНК состоит из высокоповторяющихся последовательностей, которые, как правило, не транскрибируются.
В хромосомах человека в центромерной области располагается а-сателлитная (альфоидная) ДНК, состоящая из следующих один за другим мономеров длиной 170-171 п.н. Существуют различные семейства этого типа ДНК. Одни из них специфичны для определенных хромосом, другие выявляются в нескольких хромосомах, третьи представлены несколькими субсемействами в одной хромосоме.
Основная часть перицентромерного района также относится к группе сателлитных ДНК. Сателлитные ДНК человека разделяются на две группы. Представители первой группы - это простые последовательности небольших размеров. Сателлиты этого типа называют «классическими», они расположены в прицентромерных районах хромосом. К ним относятся сателлиты 1, 2, 3. Сателлит 1 - это отрезок из 42 п.н., он обнаружен в метацентрических хромосомах 3 и 4, а также в акроцентриках. Сателлит 2 - малоконсервативный пентамерный повтор. Массивы сателлита этого типа присутствуют в хромосомах 2,10, и 16. Сателлит 3 человека — пентамерный повтор, чередующийся с декамером. Ко второй группе относятся блоки сатДНК, состоящие из отрезков 100-200 п.н. Они встраиваются в области формирования кинстохора - в центромерном районе хромосом или вплотную прилегают к нему.
Выявлены белки, взаимодействующие с сателлитной ДНК. К таковым относятся белки типа HP1 и взаимодействующие с ними белки. Предполагается, что эти белки участвуют в регуляции активности эухроматиновых генов, особенно тех из них, которые в результате хромосомных перестроек, попали в зону влияния гетерохроматина.
Тандемно повторяющиеся последовательности размером 1—4 п.н. относят к микросателлитным повторам. Наиболее часто в геноме человека встречается СА-повтор. Блоки из этих повторов можно обнаружить, примерно через каждые 30 тыс. п.н. Микросателлитные повторы используются в медицинской генетике для картирования генов различных заболеваний.
Минисателлитные повторы характеризуются большей длиной повторяющегося элемента (более 4—5 п.н.). К минисателлитным повторам формально можно отнести и теломерные повторы, однако они, в отличие от остальных, диспергированных по геному, расположены кластером и, очевидно, играют определенную структурную роль. Обычно количество теломерных повторов составляет от 250 до 1500 - общей длиной примерно 9 т.п.н. Повторы строго одинаковые, но могут различаться для разных видов.
Так, у человека основной элемент теломерных повторов представлен гексамером TTAGGG. Существует несколько семейств широко распространенных диспергированных повторов. Короткие диспергированные повторы SINE (от англ. short interspersed nucleotide element) длиной 100-500 п.н. имеют по краям короткие прямые повторы и ро1у(А)-последовательность на 3'-конце. Наиболее известный повтор этого типа - Alu-повтор. Его длина - 300 п.н., он состоит из двух мономеров и имеет ро!у(А)-хвост на 3'-конце. У человека на гаплоидный геном приходится 105—106 копий Аlu-последовательностей. Эти повторы встречаются между генами, в интронах, в сателлитной ДНК и в любой ориентации относительно друг друга. Наряду с различными SINE-семействами повторов геномы млекопитающих содержат семейство LlNE-повторов (от англ. long interspersed nucleotide element).
LINE-повтор представляет собой один из вариантов ретротранспозонов, в структуре которого присутствует ген обратной транскриптазы, но нет длинных концевых повторов LTR по флангам от кодирующей области. Единицы повтора у разных членов семейства имеют размер не более 6-7 т.п.н.; они отличаются по нуклеотидной последовательности. Частота копий различных членов семейства LINE-1 у человека измеряется от тысяч до сотен тысяч копий на геном,
Повторы в геноме человека встречаются не только в некодируюших, но и кодирующих участках некоторых генов. Интересны в этом плане цепи коллагеновых пептидов первого типа. Этот тип коллагена находится в коже, сухожилиях, костях, строме внутренних органов. Коллаген состоит из трех ct-цепей, закрученных одна вокруг другой. В основе строения цепей коллагена лежит повтор из трех аминокислот: пролина, оксипролина и глицина, способствующий плотной упаковке полипептидных цепей в фибриллах. Повторам аминокислот в полипептидных цепях коллагена соответствуют повторы кодонов в экзонах гена Colla 1. Из этого примера видно, что не только уникальные последовательности, но, и повторы в генах могут иметь функциональное значение в кодируемых ими полипептидньгх цепях.
Таким образом, роль повторяющихся элементов генома может быть различной. Некоторые из них, например теломерные и альфоидные повторы, имеют структурное значение; то же время повторяющиеся гены рРНК обеспечивают более высокий уровень синтеза продукта, т.е. играют функциональную роль. Роль микросателлитных повторов, диспергированных по геному, а также Alu, LINE и многих других повторяющихся последовательностей пока остается неясной.
- Читать далее "Хромосома и хроматин. Упаковка генетического материала в хромосоме."
Оглавление темы "Строение хромосом и генов.":1. Структура и определение гена. История изучения гена.
2. ДНК. Строение и структура ДНК. Свойства ДНК.
3. Передача генетической информации в клетке. Репликация молекулы ДНК.
4. Что такое геном? Организация генома бактерий.
5. Плазмиды бактерий. Мигрирующие генетические элементы бактерий.
6. Геном РНК вирусов. Особенности генома РНК вирусов.
7. Эукариотический геном. Повторы или избыточность генома.
8. Хромосома и хроматин. Упаковка генетического материала в хромосоме.
9. Структура хромосом. Эухромотин. Гетерохромотин.
10. Строение политенных хромосом. Структура хромосом типа ламповых щеток.