
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Плазмиды бактерий. Мигрирующие генетические элементы бактерий.
Наряду с основной, хромосомной, ДНК, бактерии могут иметь в своем составе внехромосомные ДНК-элементы, - плазмиды, обусловливающие различные свойства, например, устойчивость к антибиотикам, сульфаниламидам, способность синтезировать бактериоцин, гемолизин и т.д. При этом часть плазмид передается от одной бактериальной клетки к другой при конъюгации (так называемые конъюгативные плазмиды). Плазмиды другой группы (неконъюгативные) неспособны к конъюгативному переносу. Именно к конъюгативным относятся наиболее известные F- и R-плазмиды Escherichia coli.
Мигрирующие (мобильные) генетические элементы. К ним относятся ретротранспозоны и транспозоны, включающие и инсерционные последовательности бактерий.
Инсерционные последовательности (IS) — наиболее простые мобильные элементы. Частота их встраивания (КИ-Ю-7) сравнима с частотой спонтанных мутаций. Размеры IS-элементов относительно невелики - не более 1500 п.н. Они содержат ген транспоэазы и фланкирующие его короткие инвертированные повторы (1TR -inverted terminal repeats). При перемещении IS исходный элемент остается на прежнем месте, а реплицированная копия встраивается в новый локус. Инсерции при внедрении инактивируют кодирующие и регуляторные последовательности как в локусе-мишени, так и в соседних генах.
В IS-элементах есть кодоны-инициаторы, терминаторы, а также промоторные участки, которые могут влиять на экспрессию не только своих, но и рядом расположенных генов.
Простые транспозоны бактерий. Транспозоны этого типа в центральной части имеют для репликативной транспозиции: ген транспозазы (tnр А) и ген резольвазы (tnp R). В качестве примера мобильного элемента такого типа можно привести транс-позон Тп3 у Е. coli. Длина его центральной части составляет =5 т.п.н., концевые повторы насчитывают всего 38 п.н.
Сложные транспозоны бактерий. Такого типа мобильные элементы, помимо генов, отвечающих за транспозицию, несут дополнительные гены (резистентности к антибиотикам, сульфамидам, тяжелым металлам). Размер их генома варьирует в пределах от 2500 п,н. (в Тп9) до 9300 п.н.. На концах сложных транспозонов имеются IS-элементы с различной ориентацией: прямой - у Тn9 и обратной - у Тn10. Длина дуплицированной мишени - сегмента ДНК, в который встраиваются транспозоны, невелика: 5-12 п.н.
Механизм транспозиции бактерий. Согласно одной из моделей, вначале в реципиентной и донорной ДНК происходят одноцепочечные разрывы и их воссоединение с образованием коинтетрата при участии транспозазы. Затем следует репликация и рекомбинация, в результате чего происходит обмен участками ДНК и диссоциация коинтеграта, катализируемые резольвазой.
Транспозиция другого типа — простое встраивание - происходит без репликации. При этом фермент производит двухцепочечные ступенчатые разрезы в хромосоме-реципиенте и в мобильном элементе, а затем концы мобильного элемента присоединяются к концам хромосомы. В этом случае по краям транспозона образуются одноцепочечные фланкирующие последовательности, по матрицам которых синтезируется ДНК для заполнения бреши. Это приводит к дупликации сайта-мишени размером от 3 до 12 нуклеотидов.
Перемещение транспозонов по геному обеспечивается ферментом транспо-зазой, а их вырезание происходит в результате гомологичной рекомбинации между дуплицированными сайтами-мишенями. Транспозоны могут встраиваться в плазмиды и фаги и вместе с ними перемещаться не только между клетками внутри вида, но и между отдаленными видами бактерий.
В бактериальной клетке самостоятельно реплицируются хромосомная, фаговая и плазмидная ДНК, каждая из этих молекул представляет собой единицу репликации - репликон со своим участком инициации (ori — origin). Как оказалось, в объединении репликонов участвует транспозаза (ТпрА-белок), а их разделение осуществляет резолваза - продукт гена tnpR. В половом факторе F и в хромосомах бактерий есть копии IS-элементов (до 6 копий [S3 и I2—IS2), которые, как считают, способствуют интеграции F-фактора в бактериальную хромосому.
Встраивание и вырезание IS- и Tn-элементов вызывают различные типы перестроек в бактериальной хромосоме - делеции, инверсии, транслокации, дупликации. Так, неточное вырезание транспозона может привести к делеции соседних последовательностей. Для образования инверсий две копии мобильного элемента должны располагаться в обратной ориентации. При конъюгации этих повторов и рекомбинации между ними может произойти переворот участка хромосомы, расположенного между мобильными элементами, на 180 градусов. При неравной рекомбинации между идентичными гомологичными элементами могут образоваться дупликации и делеции в рекомбинантных хромосомах.
- Читать далее "Геном РНК вирусов. Особенности генома РНК вирусов."
Оглавление темы "Строение хромосом и генов.":1. Структура и определение гена. История изучения гена.
2. ДНК. Строение и структура ДНК. Свойства ДНК.
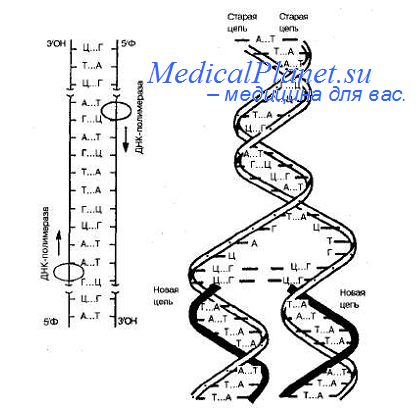
3. Передача генетической информации в клетке. Репликация молекулы ДНК.
4. Что такое геном? Организация генома бактерий.
5. Плазмиды бактерий. Мигрирующие генетические элементы бактерий.
6. Геном РНК вирусов. Особенности генома РНК вирусов.
7. Эукариотический геном. Повторы или избыточность генома.
8. Хромосома и хроматин. Упаковка генетического материала в хромосоме.
9. Структура хромосом. Эухромотин. Гетерохромотин.
10. Строение политенных хромосом. Структура хромосом типа ламповых щеток.