
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Регуляция активности желез. Клетки желез, транспортирующие ионы
Железы обычно обладают чувствительностью как к нейральным, так и к эндокринным регуляторным воздействиям. Однако часто один вид регуляции их деятельности является преобладающим.
Так, например, экзокринная секреция поджелудочной железы зависит главным образом от стимуляции гормонами секретином и холецистокинином (греч. chole — желчь + kystis — пузырь + kinein — двигать). Напротив, слюнные железы находятся преимущественно под нейральным контролем.
Нейральная и эндокринная регуляция деятельности желез обеспечивается благодаря действию химических веществ, известных как химические посредники.
Клетки желез, транспортирующие ионы
Все клетки обладают способностью транспортировать некоторые ионы против градиентов концентрации и электрических потенциалов, используя АТФ в качестве источника энергии. Этот процесс — активный транспорт — назван так, чтобы отличать его от пассивной диффузии по градиенту концентрации.
У млекопитающих концентрация иона натрия (Na+) во внеклеточной жидкости составляет 140 ммоль/л, тогда как внутриклеточная концентрация равна 5—15 ммоль/л. К тому же внутренняя часть клетки электрически отрицательно заряжена по отношению к внеклеточной среде.
В этих условиях Na+ будет стремиться проникнуть в клетку посредством диффузии как по электрическому градиенту, так и по градиенту концентрации. Клетка использует Мg2+-активируемую Na+/К+-АТФазу (натриевый насос) для удаления Na+, тем самым поддерживая требуемую низкую внутриклеточную концентрацию этого иона, расходуя при этом энергию, накопленную в виде АТФ.
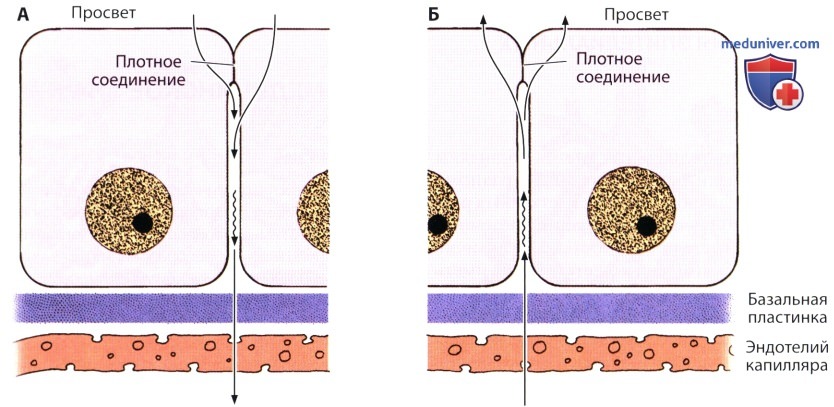
А — направление транспорта — от просвета органа к кровеносному сосуду, как это происходит, например, в желчном пузыре или кишке. Этот процесс известен как всасывание.
Б — транспорт осуществляется в противоположном направлении, как, например, в сосудистом сплетении головного мозга, цилиарном (ресничном) теле или потовой железе. Такой процесс известен как секреция. Обратите внимание на то, что для поддержания компартментализации (разделения эпителия на части — компартменты), а благодаря ей регуляции распределения ионов, требуется присутствие замыкающих соединений.
Некоторые эпителиальные клетки переносят ионы и жидкости через эпителий от апикального полюса к базальному или в обратном направлении; этот процесс известен как трансцеллюлярный перенос, или транспорт.
В кишке, проксимальных извитых канальцах почки, исчерченных протоках слюнной железы и желчном пузыре ток направлен от апикального полюса к базальному. Он осуществляется в противоположном направлении в других эпителиальных выстилках, таких, как сосудистое сплетение мозга и ресничное (цилиарное) тело.
В обоих случаях в процессе транспорта важную роль играют плотные соединения. Поскольку они перекрывают апикальные части клеток благодаря своей относительной непроницаемости для ионов, воды и более крупных молекул, они препятствуют обратной диффузии материалов, которые уже были транспортированы через эпителий. В противоположном случае происходила бы напрасная трата большого количества энергии.
Клетки проксимальных извитых почечных канальцев являются хорошим примером того, насколько организованным может быть трансцеллюлярный транспорт. Апикальная поверхность этих клеток свободно проницаема для ионов Na+, имеющихся в просвете канальца. Для поддержания электрического и осмотического баланса за ионами Na+ в клетку следуют эквимолярные количества хлорида и воды.
Базальные поверхности этих клеток содержат выраженные складки; на электронных микрофотографиях видны многочисленные длинные инвагинации (вдавления) базальной плазматической мембраны. Помимо этого, имеются обширные интердигитации базальных отростков между соседними клетками.
Показано, что Mg2+-активируемая Na+/К+-АТФаза локализована в складках как базальной плазматической мембраны, так и латеральных мембран. Между инвагинациями располагаются митохондрии, которые снабжают энергией (АТФ) механизм активного выделения Na+ из базальной части клетки во внеклеточное пространство. Хлорид и вода вновь следуют за ионом натрия пассивно. Указанным путем натрий возвращается в кровоток, а не теряется массивно с мочой.
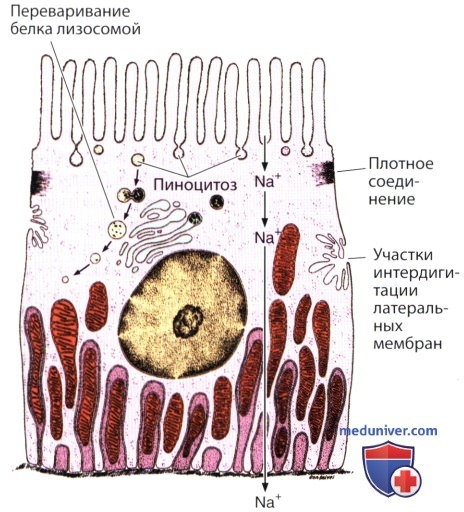
Инвагинации базальной клеточной мембраны соответствуют участкам, заполненным удлиненными митохондриями.
Такая типичная структурная организация имеется в клетках, транспортирующих ионы.
Интердигитации соседних клеток образуют структуры в виде «замков» с аналогичными выростами данной клетки.
Белок, поглощенный из просвета канальца посредством пиноцитоза, переваривается лизосомами. Ионы натрия пассивно диффундируют через апикальные мембраны этих клеток.
Далее указанные ионы активно удаляются из клеток с помощью Na+/К+-АТФазы, расположенной в их базолатеральной мембране.
Энергия для этого натриевого насоса вырабатывается расположенными рядом митохондриями.
Клетки желез, осуществляющие транспорт посредством пиноцитоза
Внеклеточные молекулы попадают в цитоплазму большинства клеток (интернализируются) с помощью пиноцитозных пузырьков, которые в изобилии формируются на плазмолемме. Этот процесс активно протекает в однослойном плоском эпителии, выстилающем кровеносные и лимфатические капилляры (эндотелий) или полости тела (мезотелий).
Клетки этого эпителия имеют небольшое количество органелл при наличии огромного числа пиноцитозных пузырьков, которые обнаруживаются на поверхностях клетки и в цитоплазме.
Эти наблюдения в сочетании с результатами, полученными путем инъекции электронно-плотных коллоидных частиц (например, ферритина, коллоидного золота, тория) с последующим исследованием препаратов под электронным микроскопом, показывают, что пузырьки, транспортирующие введенный материал, перемещаются через клетки в обоих направлениях и способны доставлять свое содержимое к поверхности клетки.
- Читать далее "Серозные клетки желез: строение, функции"
Оглавление темы "Гистология железистого эпителия":- Строение, функции стереоцилий, ресничек и жгутиков
- Типы эпителиев. Покровные эпителии
- Железистые эпителии: типы
- Физиология эпителиальных тканей. Метаплазия
- Регуляция активности желез. Клетки желез, транспортирующие ионы
- Серозные клетки желез: строение, функции
- Клетки желез секретирующие слизь. Строение, функции бокаловидной клетки
- Диффузная нейроэндокринная система (ДНЭС) - APUD: строение, функции
- Миоэпителиальные клетки желез: строение, функции
- Клетки желез секретирующие стероиды: строение, функции