
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Переход вирусного генома в клеточный геном. Паповавирусы
В случае ретровирусов показано, что уже через 1—9 ч после заражения клетки с помощью обратной транскриптазы вирусспецифическая ДНК синтезируется на матрице вирусной 70S РНК (Varmus е. а., 1975). ДНК-провирус ретровирусов имеет кольцевую форму и молекулярную массу 6х106, то есть соответствует одной субъединице вирионной РНК.
ДНК-провирус интегрирует, по-видимому, как в специфические (уникальные) участки клеточной ДНК или в тандем с эндогенным ДНК-провирусом в случае гомологичных клеток (происходит добавление вирусспецифической информации), так и в разные места клеточного генома. Клетки гетерологичных животных, в которых ДНК-провирус, родственный трансформирующему вирусу, отсутствует, вследствие интеграции заново приобретают вирусные гены.
Таким образом, в настоящее время имеются достоверные данные относительно конечного этапа перехода (интеграции) вирусспецифической информации от вирионов (вирусных частиц) к геному клетки (наличие генома опухолеродного вируса в хромосомной ДНК клеток). Однако до сих пор последовательные, промежуточные этапы этого перехода остаются невыясненными.
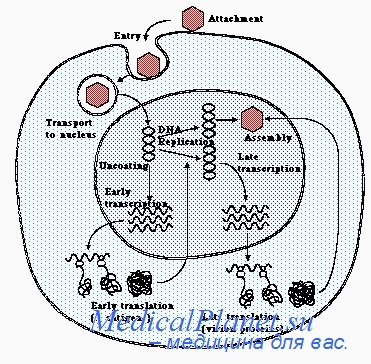
Интеграция вирусного генома с клеточной ДНК — первый этап общепринятой современной схемы вирусного онкогенеза. Следующими основными этапами, необходимыми для иеопластического превращения клеток, являются: 1) экспрессия интегрированного вирусного генома, то есть его транскрипция и трансляция с образованием вирусспецифичсских белков и 2) функционирование белка — продукта специфического вирусного онкогена, ответственного за опухолевую трансформацию. Предполагают, что этот белок нарушает регуляцию клеточного деления и поддерживает трансформированный фенотип клеток.
Итак, интеграция является центральным событием и обязательным условием опухолевой трансформации, однако не ее прямой причиной, поскольку она необходима только для включения и передачи последующим популяциям клеток вирусных онкогенов.
Паповавирусы
Основные положения вирусогенетической теории рака на протяжении 20-летнего периода ее существования способствовали исключительному прогрессу в исследованиях механизмов репродукции ДНК- и РНК-содержащих опухолеродиых вирусов, экспрессии их геномов и взаимодействия с клетками-мишенями, в некоторых случаях приводящего к опухолевой трансформации.
Сочетание молекулярно-биологических и генетических подходов (анализ с помощью различных бактериальных рестриктаз, молекулярная гибридизация, использование ts-мутантов и генетических мутантов и т. д.) позволило составить весьма полные генетические карты мелких (диаметр вириона — от 43 до 55 нм2) ДНК-содержащих онковирусов: полиомы и SV40 (Khoury е. а., 1976; Ferdinand е. а., 1977; Dhar е. а., 1978). В настоящее время эти вирусы всесторонне исследованы и стали классическими моделями вирусного канцерогенеза.
- Читать далее "Строение полиомавирусов. Геном паповавирусов"
Оглавление темы "Вирусная теория происхождения опухолей":1. Онкогенные вирусы. Молекулярно-биологические механизмы вирусного онкогенеза
2. ДНК-провирус. Интеграция вирусного генома
3. Теория протовируса. Изначальная интеграция генома вирусов
4. Геном эндогенного вируса как причина рака. Взаимодействие вирусного генома с геномом клетки
5. Варианты интеграции вирусного генома. Экспрессия интегрированных вирусных ДНК-копий
6. Место прикрепления вирусной ДНК к ДНК клетки. Места интеграции вирусного генома SV40
7. Копии вируса в геноме клетки. Геном саркомы Рауса
8. Переход вирусного генома в клеточный геном. Паповавирусы
9. Строение полиомавирусов. Геном паповавирусов
10. Ген А — онкоген полиомавирусов. Функции гена А паповавирусов