
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Вирус гриппа в неврологии - строение, вызываемые инфекции
Ортомиксовирусы (Orthomyxoviridae) семейство вирусов со спиральным капсидом и липопротеиновой оболочкой. Размер вирионов — 8-120 нм, нуклеиновая кислота — 7 фрагментов одноцепочечной линейной РНК.
К типичным представителям ортомиксовирусов в детской неврологии относятся вирусы гриппа (Influenza), являющиеся не только пневмо-, но и нейротропными.
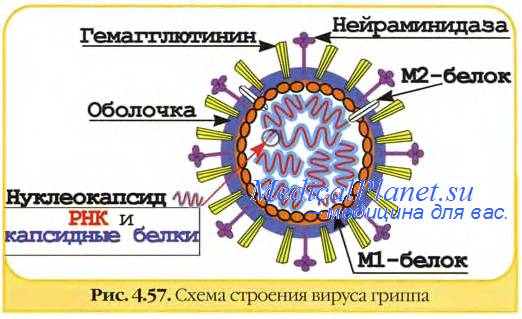
Геном вируса гриппа состоит из 8 фрагментов однонитчатой РНК, кодирующих 10 вирусных белков. Фрагменты РНК обладают общей соединяющей белковой оболочкой, образующей антигенно-стабильный рибонуклеопротеид (S-антиген). Именно S-антиген определяет принадлежность вируса к определенным серотипам (А, В или С). Снаружи вирус покрыт двойным липидным слоем, с внутренней стороны которого имеется слой мембранного белка.
Над оболочкой вируса поднимаются «шипы» двух типов (гликопротеины) — гемагглютинин и нейраминидаза. Они являются поверхностными АГ вируса гриппа и одновременно факторами его агрессии. Так, гемагглютинин определяет интенсивность интоксикации при гриппе, а нейраминидаза обладает выраженным иммуносупрессивным эффектом. Оба поверхностных АГ характеризуются выраженной способностью к изменчивости, приводя к появлению новых антигенных вариантов вируса гриппа. Гемагглютинин типов 1, 2 и 3, а также нейраминидазы типов 1 и 2 содержат вирусы, поражающие людей (включая детей первых лет жизни).
Вирус гриппа А. Более вирулентен и контагиозен, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 типа гемагглютинина (Н0, H1, Н2, НЗ).
Современный вирус гриппа A (H3N2) с трудом размножается в куриных эмбрионах, не агглютинирует эритроциты кур, взаимодействуя только с эритроцитами человека или морской свинки, имеет низкую иммуногенную активность.
Вирус гриппа В. По своей вирулентности и эпидемиологической значимости уступает вирусу гриппа А. По антигенным свойствам гемагглютинин и нейраминидазу вируса В разделяют на 5 подтипов, их изменчивость более постепенная и медленная. Вирус гриппа В содержит 1 тип гемагглютинина и 1 тип нейраминидазы, а поэтому имеет лишь антигенные варианты внутри одного серотипа.
В настоящее время в мире циркулируют дла варианта вируса гриппа В: В/Пекин/184/93 и В/Виктория/2/87. Вирусы гриппа А и В высокочувствительны к ингибиторам гемагглютинирующей и инфекционной активности (содержатся в сыворотке крови здоровых животных).
Вирус гриппа С. Вирус гриппа С содержит только гемагглютинин (не содержит нейраминидазу, но обладает рецептором, разрушающим этот энзим). Вирусу гриппа С не свойственна изменчивость. Обычно этот тип вируса не вызывает эпидемий, но может служить причиной вспышек заболевания в организованных детских коллективах (обычно приводит к легким и бессимптомным формам болезни). Антигенные и биологические свойства вируса гриппа С достаточно стабильны. Ему присущи низкая репродуктивная активность в клеточных системах и наличие рецепторов на мембране эритоцитов, отличающихся от рецепторов вирусов гриппа А и В (рецептор-деструктивная активность вируса С связана не с нейраминидазой, а с нейраминат-О-ацетилэстеразой).
Все вирусы гриппа мало устойчивы во внешней среде и под действием дезинфицирующих растворов и УФО, при нагревании до 50-60°С инактивируются в течение нескольких минут. В замороженном состоянии (при температуре -70°С) они сохраняются годами, не теряя инфекционных свойств. Оптимальное для вируса размножение происходит при температуре +37°С (в слабощелочной среде).
В настоящее время имеются доказательства возможности длительной персистенции вируса гриппа (от нескольких месяцев до пожизненной).
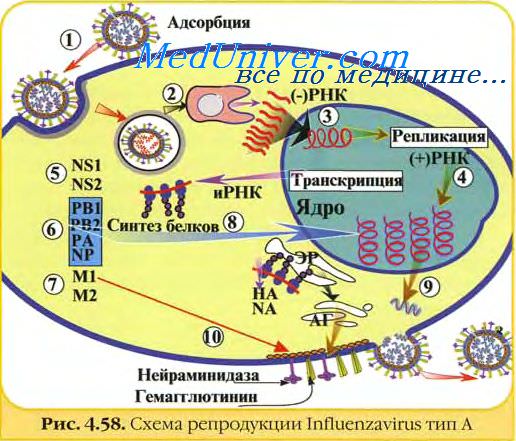
Входными воротами для вирусов гриппа служит эпителий дыхательных путей, а основной мишенью — клетки цилиндрического реснитчатого эпителия. Первой линией иммунной защиты от вирусов гриппа являются неспецифические факторы (вязкие свойства слизи, движение ресничек цилиндрического эпителия, ингибиторы репликации вируса в секрете дыхательных путей, макрофаги, секреторный IgA).
Роль дополнительных факторов защиты также выполняют конагглютинин, белки сурфактанта (А и D), манозосвязывающий белок. Все они являются лектинами С-типа, оказывающими опсонизирующее действие вследствие связывания с углеводами вируса и индукцией его агрегации.
Попав в носоглотку, вирус гриппа подвергается действию секрета клеток, белки которых неспецифически ингибируют гемагглютинирующую и инфекционную вирусную активность (в основном за счет содержания в секреторных гликопротеидах большого количества N-ацетилнейраминовой кислоты). Погибшие клетки элиминируются макрофагами и удаляются с секретом респираторного тракта.
В подслизистом слое происходит высвобождение таких биологически активных веществ (БАВ), как серотонин, гистамин, простагландины и кинины, которые вызывают ряд патогенетически значимых феноменов: смещение рН в «кислую» сторону, сосудистые реакции, отек, местные нарушения метаболизма, реакция форменных элементов крови, образование мелких тромбов и кровоизлияний, диапедез мононуклеаров и нейтрофильных лейкоцитов и др., с максимальной выраженностью воспалительного процесса в трахее и бронхах.
Нейротоксикоз при гриппе является прямым следствием нейротропности вируса, резорбции токсичных субстанций вируса, а также продуктов нарушенного метаболизма. Вирус гриппа оказывает токсическое влияние на сосудистую систему, вызывая нарушения микроциркуляции и развитие геморрагического синдрома. Вирусемия сопровождается фазовыми повреждениями симпатического и парасимпатического отделов вегетативной нервной системы.
Вторая линия специфической зашиты от вируса гриппа представлена в более глубоко расположенных слоях эпителия. Это ИФН, циркулирующие антитела классов IgM, IgG и IgE. При инфицировании вирусом гриппа развиваются ранние цитокиновые реакции, которые могут представлять естественный вариант реагирования на вирус, когда сам патоген индуцирует систему ИФН.
После внедрения вируса и индукции образования ИФН происходит деструкция информационных РНК вируса гриппа под воздействием 2'-5'-олигоаденилсинтетазы и активированной эндонуклеазы. Одновременно фосфорилирование а-субъединицы фактора, инициирующего трансляцию активированной протеинкиназой, блокирует синтез вирусных белков. Совокупность описываемых процессов приводит к угнетению репродукции широкого спектра РНК и ДНК-содержащих вирусов за счет прямого антивирусного эффекта (внутриклеточного).
ИФН-b активирует NK-клетки и цитотоксические лимфоциты, защищая окружающие незараженные клетки от инфицирования. Он связывается со специфическими рецепторами здоровых неповрежденных клеток, превращая их в невосприимчивые к вирусу. ИФН-b активирует почти все иммунные реакции (модуляция взаимосвязи эндокринной и иммунной систем, активация макрофагов, повышение цитотоксичности, стимуляция экспрессии антигенов HLA классов I и II).
Когда описанных выше эффектов ИФН недостаточно для подавления активности вирусной инфекции, развивается острое заболевание. Оно сопровождается продукцией каскада ранних цитокинов, активацией CD4+ и CD8+ Т-лимфоцитов с последующим развитием специфического Т-и В-опосредованного клеточного иммунитета. При этом (помимо ИФН) происходит синтез ранних цитокинов ФНО, ИЛ-1, ИЛ-6, ИЛ-10, ИЛ-15, трансформирующего фактора роста (ТФР), но именно продукция ИФН-b рассматривается в качестве ключевого признака инфицирования вирусом.
ИФН-а и ИФН-b вызывают перераспределение клеток для последующих иммунорегуляторных эффектов, они также могут усиливать при гриппе специфический иммунный ответ в периоде первичной инфекции, привлекая Т- и В-клеточные популяции в места локального воспаления для представления антигена.
Макрофаги первыми реагируют на вирус гриппа, расщепляя его на отдельные пептиды. Последние, оказавшись на мембране макрофага, вступают в контакт с молекулами HLA классов I и II.
ИФН, секретируемый ИЛ-1, усиливает экспрессию антигена комплекса HLA. Синергичное действие ИЛ-1 и ИЛ-2 способствует пролиферации предшественников Т-клеток в тимусе, в очаг инфекции направляются Т-хелперы (CD4+), которым представляются фрагменты антигена в комплексе с молекулами HLA. В активации CD4+ Т-лимфоцитов участвуют ИЛ-4, ИЛ-6 и ФНО.
Возможно непосредственное связывание вирусных антигенов Т-лимфоцитами при участии антигена комплекса HLA класса П. Затем к этому процессу подключаются CD8+ (цитотокси-ческие Т-лимфоциты) и NK-клетки. CD8+ клетки могут как элиминировать вирус из нижних отделов дыхательных путей, так и усиливать реакции локального воспаления.
При неэффективной защите макроорганизма от вируса гриппа факторами клеточного иммунитета в процесс включается гуморальный иммунитет (местный и общий). Секреторный IgA (SIgA) — основа специфического местного иммунитета. SIgA синтезируется клетками реснитчатого и секреторного эпителия, макрофагами и Т-лимфоцитами. Макрофаги захватывают обломки разрушенных вирусом клеток и метаболизируют их, а антигены, подготовленные макрофагами, активизируют Т- и В-лимфоциты, которые, достигнув стадии плазмоцитов, также продуцируют антитела класса IgA. Последний выполняет эффекторные функции и играет роль молекулы, регулирующей функции клеток иммунной системы (например, альвеолярных макрофагов с рецепторами к Fc-фрагменту IgA). Иммунные комплексы (ИК), содержащие антитела IgA, индуцируют в моноцитах-макрофагах синтез ФНО-а и СЗ-компонента комплемента. SIgA связывают вирус и препятствуют его выходу из организма в активной форме.
Антитела IgM формируются через 3-5 дней после встречи с вирусом, а их повышенное содержание свидетельствует об остроте инфекционного процесса. Иммунные механизмы при первичной встрече с вирусом гриппа запоминаются клетками памяти.
В окружении антител вирус гриппа может сохранять свою структуру, а при разрушении иммунного комплекса (антиген-антитело) вновь поражать чувствительные клетки; длительная циркуляция ИК приводит к инфицированию все большего количества клеток и к персистенции вируса.
При массивной виремии происходит токсическое поражение сосудистого аппарата ЦНС; отмечается гиперсекреция спинномозговой жидкости и нарушения ликвородинамики (повышение внутричерепного давления, в тяжелых случаях — отек мозга). Чаще повреждаются мягкие оболочки мозга и сосудистое сплетение, где выявляются антигены вируса гриппа.
Устойчивость организма новорожденного к вирусу гриппа может определяться наследственной резистентностью. Приобретенный иммунитет появляется только после перенесения болезни.
- Читать "Аденовирусы (Adenoviridae) в неврологии - строение, вызываемые инфекции"
Оглавление темы "Вирусные инфекции в неврологии":- Грипп у новорожденных детей - причины, диагностика, лечение
- Патогенез (механизмы развития) вирусных нейроинфекций
- Виды (представители) нейротропных вирусов
- Герпесвирусы (Herpesviridae) - строение, вызываемые инфекции
- Энтеровирусы (Enteroviridae) в неврологии - строение, вызываемые инфекции
- Вирус кори (Morbilli) в неврологии - строение, вызываемые инфекции
- Вирус паротита в неврологии - строение, вызываемые инфекции
- Вирус краснухи (Rubivirus) в неврологии - строение, вызываемые инфекции
- Вирус гриппа в неврологии - строение, вызываемые инфекции
- Аденовирусы (Adenoviridae) в неврологии - строение, вызываемые инфекции