
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Патогенез спиноцеребеллярных атаксий - СЦА8, СЦА10 и СЦА12
Особый патогенез имеют формы аутосомно-доминантных спиноцеребеллярных атаксий, обусловленные экспансией повторов (CTG)n, (АТТСТ)n и (CAG)n в некодирующих областях генов -СЦА8, СЦА10 и СЦА12.
В настоящее время предполагается, что для этих заболеваний характерен принципиально новый молекулярный механизм нейродегенерации, связанный с токсическим эффектом мутантной мРНК (Ranum L.P., Day J.W.).
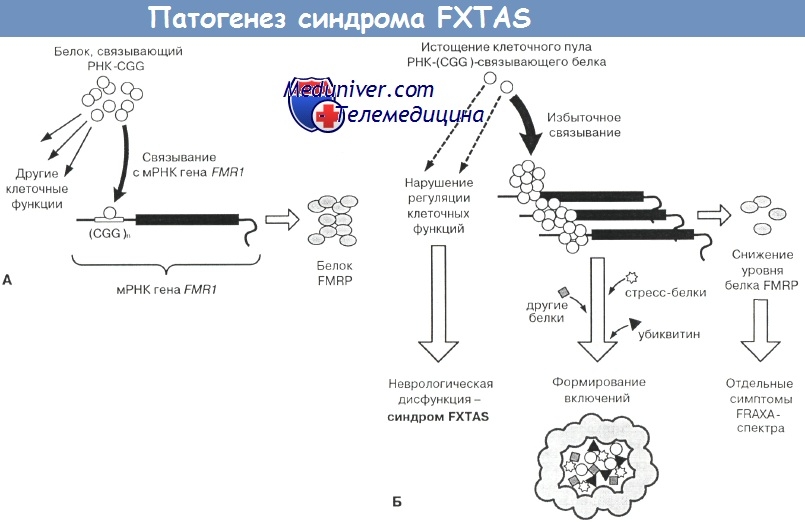
К этому же классу патологии относятся и две другие болезни экспансии — миотоническая дистрофия и недавно описанная новая форма дегенеративной атаксии, получившая обозначение «синдром FXTAS» (Hagerman P.J., Hagerman R.J.).
Подробно механизм «РНК-токсической» модели нейродегенерации представлен в статье, посвященном синдрому FXTAS. Сущность такого механизма заключается в том, что имеющиеся в увеличенном числе молекулы мутантной мРНК с удлиненным три(пснта-)-нуклеотидным повтором становятся объектом избыточного взаимодействия с особыми регуляторными РНК-связывающими белками.
В результате этого нормальный пул данных белков в клетке истощается, что сопровождается нарушением их регуляторных функций и дегенерацией определенных групп нейронов. Параллельно этому избыточные РНК-белковые комплексы становятся «ядром» полимеризации для других пептидных молекул (убиквитина, белков теплового шока и т.д.); это закономерно приводит к формированию разнообразных по составу мультибелковых нейрональных включений — маркеров различных форм нейродсгснсративных атаксий (Ranum L.P., Day J.W., Hagerman P.J., Hagerman R.J.).
Развитие самостоятельной формы аутосомно-доминантной атаксии может быть связано также с нарушением сигнального фосфорилирования клеточных субстратов, как это показано для CUA14, являющейся результатом мутаций в гене протсинкиназы С-у (PRKCG) (Chen D.H. et al.).
Белок PRKCG является представителем семейства серин-треониновых киназ и имеет высокий уровень экспрессии в клетках Пуркинье. Осуществляя постоянную челночную транслокацию между плазматической мембраной и цитозолем, он играет важную роль в передаче клеточных сигналов, синаптической трансмиссии, клеточной пролиферации и дифференцировке, туморогенезе.
Доминантный эффект мутаций PRKCG у больных СЦА14 объясняют конкуренцией мутантного и нормального белков за присоединение к их мембранным сайтам или за связывание специфических субстратов. Предполагается также, что мутации могут оказывать доминантный токсический эффект посредством повышения внутриклеточной концентрации Zn2+ (следствие нарушенного связывания мутантного белка с этим ионом).
Интересно отметить, что у трансгенных ATXN1-мышей наблюдается нарушение активности и локализации PRKCG в клетках Пуркинье (Skinner P.J. et al.), что может указывать на значение белка PRKCG в развитии и прогрессировании СЦА1. С другой стороны, в клетках Пуркинье у PRKCG-мутантных пациентов с СЦА14 наблюдается снижение экспрессии атаксина-1 (продукта гена ATXN1) (Chen D.H. et al.). Эти факты позволяют предполагать наличие определенных общих путей нейродегенерации при СЦА14 и полиглутаминовых формах типа СЦА1.
Наконец, еще один самостоятельный молекулярный механизм доминантных атаксий предполагает нарушение поддержания морфофункциональной целостности нейронов в результате дефекта их трофики. Так, при СЦА27 имеет место повреждение гена фактора роста фибробластов-14 (FGF-I4) — одного из 23 известных представителей семейства факторов роста фибробластов (van Swieten J.С. et al.). Белок FGF-14 локализован в аксональных проекциях и предположительно участвует в функционировании аксонов, синаптосом и/или процессах нейротрансмиссии.
Нельзя исключить, что генетические повреждения и других факторов роста могут быть связаны с атактическими либо другими формами нейродегенерации.
а — нормальный аллель гена FMR1; б - «премутантный» аллель гена FMR1.
- Читать "Трансгенные модели аутосомно-доминантных атаксий"
Оглавление темы "Аутосомно-доминантные атаксии":- Аутосомно-доминантные атаксии - история изучения
- Классификация аутосомно-доминантных атаксий
- Генетика аутосомно-доминантных атаксий
- Эпидемиология аутосомно-доминантных атаксий - распространенность, частота
- Патогенез полиглутаминовых форм аутосомно-доминантных атаксий
- Патогенез спиноцеребеллярных атаксий - СЦА8, СЦА10 и СЦА12
- Трансгенные модели аутосомно-доминантных атаксий
- Клиника аутосомно-доминантных атаксий и их течение
- Пример аутосомно-доминантной атаксии
- Патоморфология аутосомно-доминантной атаксии - гистология