
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Нарушение распознавания клеток и клеточных сигналов L1-CAM при НСП
Разнообразие идентифицированных генов и кодируемых ими белков говорит о множественных молекулярных механизмах, вызывающих дегенерацию пирамидных путей (при поздних формах) или нарушение их развития (при ранней и врожденной НСП).
Поначалу между механизмами отдельных вариантов НСП не прослеживалось никакой общности, но последние достижения в идентификации генов и их белковых продуктов уже вполне позволяют объединить в общие группы формы со сходными патогенетическими звеньями (Casari С, Rugarli Е., Crosby A., Proukakis С., Fortini D. et al., Reid E.).
Так возникает новая классификация наследственных спастических параплегий (НСП), основанная на функциональной геномике. В этом разделе мы кратко суммируем данные о генах НСП именно с этой, функциональной, точки зрения.
E. Rcid выделяет следующие «точки приложения» известных генов и белков наследственных спастических параплегий (НСП):
1. Нарушение распознавания клеток и межклеточных контактов - L1-САМ.
2. Нарушение процессов миелинизации — PLP1.
3. Нарушения на уровне митохондриальных шаперонов — параплегии и HSP60.
4. Нарушения клеточного аксонального транспорта — KIF5A, спастин, атластин, спартин и др.
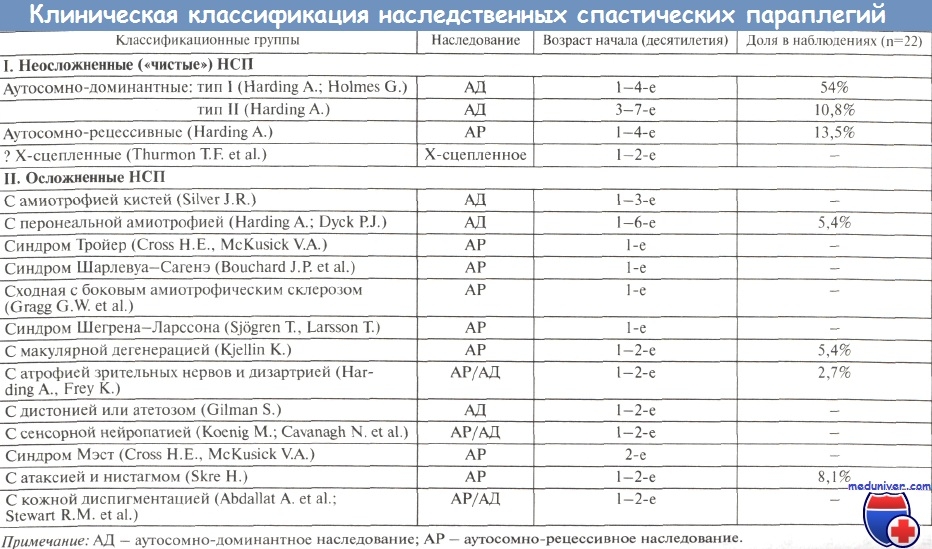
Нарушение распознавания клеток и клеточных сигналов L1-CAM
Широко экспрессирующийся в нервной системе ген L1-CAM, ответственный за форму SPG1, кодирует гликопротеин клеточной поверхности, обеспечивающий созревание, миграцию нейронов и формирование их аксональных контактов с другими нейронами, клетками глии и мышцами в процессе онтогенеза либо после травмы.
Патоморфологичсские данные и исследования трансгенных животных указывают на важную роль этого белка в нормальном развитии кортикоспинальных трактов (Casari G., Rugarli Е.), а недавно также показана его роль в обеспечении нейрональной миграции (Thelen К. et al.). Наконец, исследованиями in vitro показано участие L1-CAM в ветвлении аксонов и процессах роста нервных окончаний (Reid Е.).
В реализации многочисленных функций L1-CAM основное значение придается его межмолекулярным взаимодействиям как вне, так и внутри клетки.
Показано, что L1-CAM связывается с многочисленными внеклеточными лигандами, в частности, другими белками клеточной адгезии, интегринами, белками внеклеточного матрикса и протеогликанами. Исключительное значение имеет также связывание молекул L1-CAM друг с другом (т.е. гомофильное связывание), которое как раз и является ключевым сигнальным взаимодействием, способствующим клеточной адгезии и аксональному спрутингу.
Если говорить о внутриклеточной передаче сигналов, то L1-CAM участвует в ней разными путями. Главный из них — взаимодействие с рецепторами фактора роста фибробластов (FGFR) - мембраносвязанными белками, имеющими внутриклеточные домены, которые регулируют активность тирозинкиназ.
Гомофильное связывание L1-CAM активирует FGFR-зависимые тирозинкиназы, что, в свою очередь, усиливает приток ионов кальция в клетку и тем самым изменяет — через неизвестные пока механизмы — направление аксонального роста. Важно также взаимодействие L1-CAM (его цитоплазматического домена) с анкирином — белком, связывающим цитоскелетный белок спектрин. Благодаря этому взаимодействию гомофильное связывание L1-CAM способно перестраивать цитоскслет нейронов в процессе их роста, постепенного видоизменения их морфологии и формирования нейрональной транспортной сети по всей длине аксона.
Экспериментальные модели помогают понять патогенез формы SPG1: у мышей с нокаутным геном L1-CAM, фенотип которых характеризуется спастичностью, повреждение кортикоспинальных путей связано с отсутствием их перекреста в продолговатом мозге из-за нарушенной направленности аксонов (Dahme М. et al.).
Таким образом, патология L1-CAM и связанная с ним форма SPG1 демонстрируют наиболее «ранний» уровень повреждения имеющихся сложных механизмов формирования пирамидных путей -уровень непосредственно аксонального роста и его направленности в онтогенезе. Как показывает опыт изучения L1-CAM, эти процессы регулируются большим числом межмолекулярных и межклеточных взаимодействий, нарушение которых может сопровождаться развитием спастичности.
- Читать "Нарушение процессов миелинизации при НСП: протеолипидный белок и его изоформы"
Оглавление темы "Наследственные спастические параплегии (НСП)":- Спастическая параплегия 1-го типа (SPG1) - клиника, диагностика
- Спастическая параплегия 2-го типа (SPG2) - клиника, диагностика
- Спастическая параплегия 16-го типа (SPG16) - клиника, диагностика
- Синдром Шегрена-Ларссона - клиника, диагностика
- Пример синдрома Шегрена-Ларссона
- Спастическая атаксия типа Шарлевуа-Сагенэ - клиника, диагностика
- Ранний восходящий спастический паралич и ювенильный первичный боковой склероз - клиника, диагностика
- Нарушение распознавания клеток и клеточных сигналов L1-CAM при НСП
- Нарушение процессов миелинизации при НСП: протеолипидный белок и его изоформы
- Нарушения в системе митохондриальных молекулярных шаперонов при НСП и HSP60