
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Диагностика атаксии Фридрейха путем оценки митохондриальных нарушений
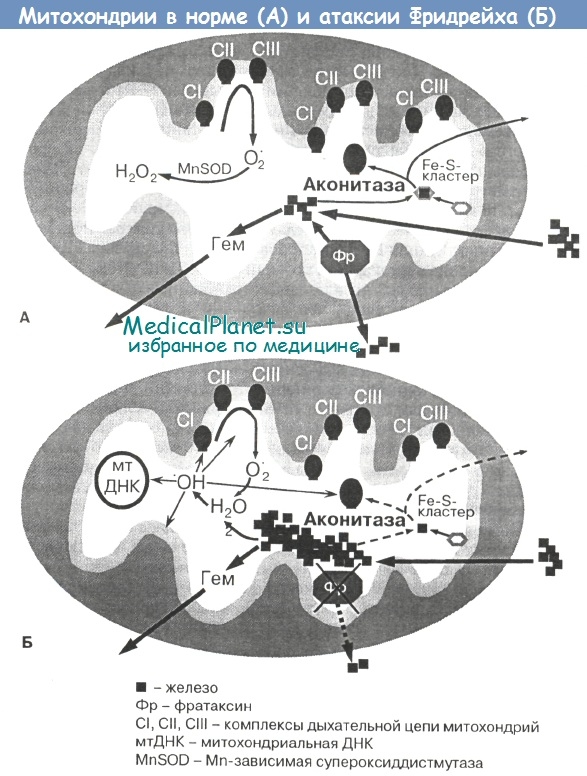
Как было показано выше, патогенетической основой атаксии Фридрейха являются нарушения синтеза АТФ в клетке и митохондриального энергетического обмена. Снижение уровня митохондриального белка фратаксина непосредственно приводит к развитию окислительного стресса и клеточной гибели, поэтому атаксия Фридрейха является уникальной моделью для большого числа нейродегенеративных заболеваний, в патофизиологии которых центральное место занимают активные формы кислорода и окислительный стресс.
Однако, несмотря на активное изучение атаксии Фридрейха с использованием самых современных технологий, конкретные дефекты энергетического метаболизма при этом заболевании остаются неизвестными. Недостаточно разработаны также прижизненные лабораторные методы оценки митохондриальной функции и адекватного биохимического мониторинга за состоянием больных.
Всестороннее изучение состояния энергообеспечения при атаксии Фридрейха может помочь в выяснении интимных механизмов патогенеза болезни, а коррекция нарушений энергообмена должна стать важным звеном лечебной тактики при данном заболевании. Кроме того, определение совокупности показателей энергообеспечения клетки может быть использовано в качестве важного вспомогательного прогностического теста.
Поскольку главным признаком митохондриальной недостаточности является нарушение активности окислительно-восстановительных ферментов, их исследование наиболее значимо как в теоретическом, так и в практическом отношении.
В настоящее время морфологический анализ биоптатов скелетных мышц служит общепризнанным, базисным тестом на наличие митохондриальной патологии. Однако в силу технической сложности и травматичности (биопсия мышцы) его использование в широкой практике для лабораторного контроля за динамикой состояния больных и оценки клинического эффекта комплексной терапии митохондриальных нарушений крайне затруднительно.
При изучении биоэнергетических процессов широко используются методы, основанные либо на измерении активности дыхательной цепи митохондрий в целом (по поглощению митохондриями кислорода), либо на определении активности отдельных ферментов дегидрогеназ. Перспективно использование для этой цели цитохимического анализа ферментного статуса лимфоцитов (Нарциссов Р.П., Петричук С.В. и др.). Показано, что активность окислительно-восстановительных ферментов клеток периферической крови, оцениваемая количественным цитохимическим методом, может адекватно отражать энергетический обмен клеток и тканей всего организма, т.е. служить своеобразным «цитохимическим зеркалом тканей» (Комиссарова И.А.); это тем более справедливо для атаксии Фридрейха, поскольку при ней указанные нарушения носят системный характер.
Доказательством адекватности применения цитохимического морфометрического анализа лимфоцитов при митохондриальных болезнях является установленная корреляция функциональной активности митохондрий лимфоцитов с общепринятыми показателями митохондриальной патологии в биоптатах мышц, в том числе с феноменом «рваных красных волокон» — феномен RRF (Сухорукое B.C. и др.). Можно заключить, что количественный цитохимический метод оценки ферментного статуса лимфоцитов периферической крови не менее достоверно, чем характеристики митохондрий в скелетных мышцах, отражает полисистемную митохондриальную недостаточность.
Впервые визуальный подсчет гранул продукта реакции для количественной гистохимии ферментов был предложен Э.Пирсом в 1962 г. В дальнейшем была предложена существенная модификация данного метода (Нарциссов Р.П.). Ферментативная активность выражается в условных единицах, соответствующих среднему числу гранул формазана — продукта цитохимической реакции. В настоящее время в практику все шире входят наиболее современные модификации метода Р.П.Нарциссова с использованием компьютерной морфометрии и специальных пакетов программного обеспечения.
При этом оцениваются количество гранул митохондриальных ферментов и их качественные параметры: площадь, форма, интервал яркости, оптическая плотность, интервал оптической плотности и др.
При атаксии Фридрейха для оценки митохондриальных нарушений с помощью цитохимического метода наиболее целесообразным представляется определение активности ряда ферментов-дегидрогеназ лимфоцитов: сукцинатдегидрогеназы (СДГ), а-глицерофосфатдегидрогеназы (ГФДГ) глутаматдегидрогеназы (ГДГ), лактатдегидрогеназы (ЛДГ), малатдегидрогеназы (МДГ) и др. Эти ферменты занимают ключевые позиции в аэробном и анаэробном энергообеспечении клетки. Так, СДГ считается классическим маркерным ферментом митохондрий, прочно связанным с их внутренней мембраной и отражающим функциональное состояние этих органелл: он катализирует одну из реакций в цикле трикарбоновых кислот Кребса и входит в состав комплекса 11 дыхательной цепи митохондрий (Ленинджер А., Berg J.M. et al.).
Фермент ГФДГ отражает челночный механизм между гликолизом и циклом Кребса, сущность которого состоит в переносе восстановительных эквивалентов от цитозольного NADH в митохондрии (Berg J.M. et al.). Снижение активности СДГ при увеличении активности ГФДГ, приводящее к резкому изменению их соотношения, может свидетельствовать о глубоком нарушении энергетического обмена в клетках и являться показателем накопления недоокислснных продуктов обмена, снижения буферных оснований крови и переключения тканевых систем на резервный путь обмена. Наряду с глицерофосфатным челночным механизмом существует малатный челночный механизм, составляющими компонентами которого являются цитоплазматическая и митохондриальная МДГ (Ленинджер А., Berg J.M. et al.).
Внутренняя митохондриальная мембрана непроницаема для NADH, поэтому перенос восстановительных эквивалентов из цитозоля в митохондрии возможен с помощью малат-аспартатного Н+-транспортирующего шунтового механизма. ЛДГ является ключевым ферментом, действующим на последнем этапе гликолиза, происходящем в анаэробных условиях и сопровождающемся восстановлением пирувата и лактата с высвобождением НАД+. Наконец, еще один исследуемый фермент — ГДГ — локализуется исключительно в матриксе митохондрий и характеризует уровень белкового обмена. ГДГ катализирует окислительное дезаминированис глутамата с образованием а-кетоглутарата, который может быть использован в цикле лимонной кислоты.
Таким образом, изменение активности указанных ферментов и нарушение их соотношения служат важным маркером митохондриальной дисфункции. Разработка и применение информативных цито- и биохимических методов контроля при болезнях клеточной энергетики необходимы для решения ряда диагностических проблем, мониторинга течения заболевания, оценки системных нарушений метаболизма в тканях и уточнения патогенетических основ заболевания.
Применительно к атаксии Фридрейха решение этих задач имеет важное теоретическое и практическое значение, учитывая развитие этого заболевания в детском или молодом возрасте, относительно высокую его распространенность и тяжелое инвалидизируюшее течение.
- Читать "Цитохимический анализ активности митохондриальных ферментов при атаксии Фридрейха"
Оглавление темы "Атаксия Фридрейха":- Лечение и профилактика врожденных мозжечковых атаксий
- Атаксия Фридрейха - история изучения
- Генетика атаксии Фридрейха
- Клинико-генетические корреляции при атаксии Фридрейха
- Фратаксин - функции, значение. Патогенез атаксии Фридрейха
- Роль патологии митохондрий в патогенезе атаксии Фридрейха
- Роль нарушений обмена аминокислот в патогенезе атаксии Фридрейха
- Диагностика атаксии Фридрейха путем оценки митохондриальных нарушений
- Цитохимический анализ активности митохондриальных ферментов при атаксии Фридрейха
- Морфометрический анализ активности митохондриальных ферментов при атаксии Фридрейха