
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизмы генных мутаций. Молекулярный генез генных мутаций.
В предыдущем разделе уже говорилось о том, что генные мутации могут представлять собой замены оснований, а также их вставки, перемещения или выпадения. Различают два типа замен оснований:
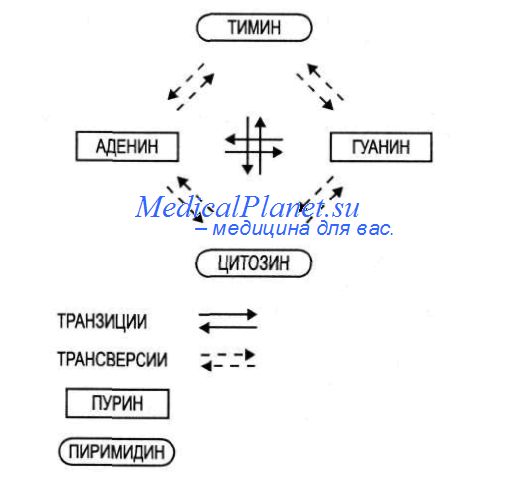
1) замены пурина на другой пурин или пиримидина на пиримидин. Их называют транзитами: А = G, Т = С (т.е. такие замены пар нуклеотидов, которые не изменяют ориентации: AT = GC; ТА = CG);
2) замены пурина на пиримидин или пиримидина на пурин. Их называют трансверсиями: А = Т, А = С, G = С, G = Т (т.е. такие замены пар нуклеотидов, которые изменяют ориентацию: AT = ТА, AT = CG).
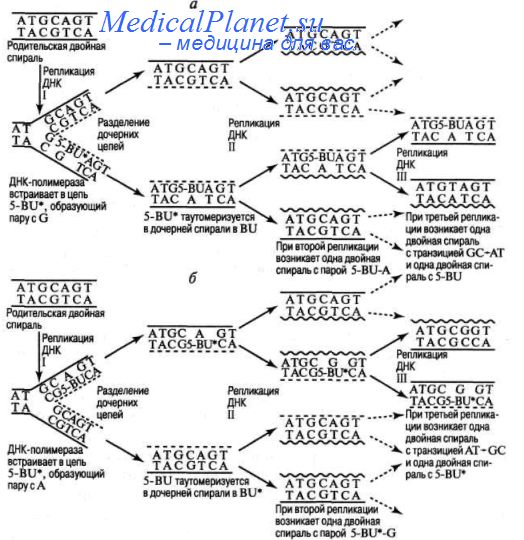
Установлено, что спонтанные транзииии могут происходить при репликации ДНК вследствие таутомеризации, изменяющей способность нуклеотидов образовывать водородные связи: аденин приобретает свойства гуанина, гуанин — аденина, цитозин - тимина, тимин - цитозина. Участие процесса репликации в мутагенезе было установлено при изучении биологических эффектов аналогов оснований ДНК, например, 5-бромурацила, вызывающего мутации у бактериофагов и бактерий.
При включении аналога в ДНК возможна ошибка, состоящая в том, что 5-бромурацил, находясь в редкой енольной форме, спаривается с гуанином, а затем в обычной кетоформе спаривается с аденином; в третьем цикле репликации аденин нормально спаривается с тимином, завершая транзицию GC — AT .
Ошибка при считывании связана с тем, что 5-бромурацил включается при репликации на место тимина, а затем спаривается, находясь в редкой енольной форме, с гуанином; в третьем цикле репликации гуанин нормально спаривается с цитозином, завершая транзицию AT - CG.
Аналогичным образом действует другой аналог оснований ДНК - 2-аминопурин. Транзииии строго в направлении GC - AT можно получить с помощью гидроксиламина, который, специфически реагируя с цитозином, переводит его в форму, способную образовывать водородные связи с аденином.
Показано, что аллели генов, контролирующих репликацию (кодирующих ДНК-полимеразу, ДНК-лигазу, ДНК-связывающие белки), проявляют мутаторную или антимутаторную активность, соответственно повышая или понижая частоту мутаций. А соотношение полимеразная/экзонуклеазная активность, как правило, уменьшается в направлении мутатор > дикий тип > антимутатор. Эти результаты согласуются с обнаружением корректирующей 3'-5'-экзонуклеазной активности у всех ДНК-полимераз, в результате которой происходит отщепление ошибочно включенных в ДНК оснований.
Следовательно, частота ошибок репликации определяется соотношением полимеразной и экзонуклеазной активности ДНК-полимеразы.
Что касается молекулярного механизма возникновения вставок, то он, возможно, связан с локальной диссоциацией двойной спирали и последующим ошибочным спариванием нуклеотидов при ее восстановлении.
Несколько особняком стоит класс так называемых динамических мутаций, или мутаций экспансии. Экспансия (расширение, увеличение) вданном случае - это превышение допустимого числа повторяющихся элементов в микро-/мини-сателлитных последовательностях, локализованных в кодирующих или регуляторных областях гена. Появление мутаций экспансии связано с нестабильностью числа тринуклеотидных повторов в транскрибируемых или регуляторных областях генов. Мутации такого типа зарегистрированы только у человека. Многие из тринуклеотидных повторов в количественном отношении характеризуются высоким уровнем популяционной изменчивости, в пределах которого не наблюдается фенотипических нарушений. Однако если их число превысит определенный критический уровень, развиваются заболевания, для которых характерны: неполное доминирование признака, геном ной кетоформе спаривается с аденином; п третьем цикле репликации аденин нормально спаривается с тимином, завершая транзицию GC — AT.
Ошибка при считывании связана с тем, что 5-бромурацил включается при репликации на место тимина, а затем спаривается, находясь в редкой енольной форме, с гуанином; в третьем цикле репликации гуанин нормально спаривается с цитозином, завершая транзицию AT-CG.
Аналогичным образом действует другой аналог оснований ДНК - 2-аминопурин. Транзиции строго в направлении GC - AT можно получить с помощью гидроксиламина, который, специфически реагируя с цитозином, переводит его в форму, способную образовывать водородные связи с аденином.
Показано, что аллели генов, контролирующих репликацию (кодирующих ДНК-полимеразу, ДНК-лигазу, ДНК-связывающие белки), проявляют мутаторную или антимутаторную активность, соответственно повышая или понижая частоту мутаций. А соотношение полимеразная/экзонуклеазная активность, как правило, уменьшается в направлении мутатор > дикий тип > антимутатор. Эти результаты согласуются с обнаружением корректирующей 3'-5'-экзонуклеазной активности у всех ДНК-полимераз, в результате которой происходит отщепление ошибочно включенных в ДНК оснований. Следовательно, частота ошибок репликации определяется соотношением полимеразной и экзонуклеазной активности ДНК-полимеразы.
Что касается молекулярного механизма возникновения вставок, то он, возможно, связан с локальной диссоциацией двойной спирали и последующим ошибочным спариванием нуклеотидов при ее восстановлении.
Несколько особняком стоит класс так называемых динамических мутаций, или мутаций экспансии. Экспансия (расширение, увеличение) вданном случае - это превышение допустимого числа повторяющихся элементов в микро-/мини-сателлитных последовательностях, локализованных в кодирующих или регуляторных областях гена. Появление мутаций экспансии связано с нестабильностью числа тринуклеотидных повторов в транскрибируемых или регуляторных областях генов. Мутации такого типа зарегистрированы только у человека. Многие из тринуклеотидных повторов в количественном отношении характеризуются высоким уровнем популяционной изменчивости, в пределах которого не наблюдается фенотипических нарушений. Однако если их число превысит определенный критический уровень, развиваются заболевания, для которых характерны; неполное доминирование признака, геном ный импринтинг и нарастание тяжести клинических проявлений в последующих поколениях (так называемая антиципация). Подробно этот материал освещается в части П. Медицинская генетика, поэтому в данной главе ограничимся лишь краткими, предварительными сведениями. Список болезней экспансии, вызванных динамическими мутациями, превысил десяток нозологических форм и продолжает постоянно пополняться.
Для части из них (синдром ломкой Х-хромосомы, миотоническая дистрофия) типична передача по материнской линии. Другие, как, например, хорея Гентингтона, передаются преимущественно по отцовской линии. Этиология этих заболеваний связана с экспансией триплетных повторов (CGG)n, (CTG)n, (CAG)n. Практически для всех «болезней экспансии» характерно поражение подкорковых структур головного мозга, причем тяжесть заболевания и его начало положительно коррелируют с числом повторов. Причиной повреждающего действия одних динамических мутаций является утрата функции вследствие блокирования генной экспрессии, других - появление белковых продуктов, аномальная структура которых оказывает повреждающее действие на жизнеспособность или функционирование клеток.
Еще один, класс мутаций — это мутации, возникающие вследствие инсерций (вставок) мобильных генетических элементов (МГЭ). Они описаны в нашей статье. Заметим только, что есть существенное различие между индукцией точковых мутаций и инсерций. При обработке мутагенами частоту точковых мутаций можно увеличить. А результаты перемещения МГЭ от действия мутагенов не зависят.
- Читать далее "Обратные мутации. Супрессоры мутаций."
Оглавление темы "Мутации у человека.":1. Генные мутации. Характеристика генных мутаций.
2. Механизмы генных мутаций. Молекулярный генез генных мутаций.
3. Обратные мутации. Супрессоры мутаций.
4. Причины мутаций. Роль ионизирующего излучения в развитии мутаций.
5. Доза ионизирующего излучения и частота мутации. Зависимость дозы ионизирующего излучения от частоты мутаций.
6. Ультрафиолетовые лучи и мутации. Воздействие ультрафиолетовых лучей на гены.
7. Роль химических соединений в развитии мутаций. Химия и мутации генов.
8. Алкилирующие соединения и мутации. Виды алкилирующих химических веществ.
9. Спонтанные мутации. Причины возникновения спонтанных мутаций.
10. Учет спонтанных мутаций у человека. Техника учета спонтанных мутаций.