
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Компенсация дозы генов у млекопитающих. Современная теория инактивации Х-хромосомы.
Еще в 1949 г. М. Барр и Ч. Бертрам обнаружили в соматических клетках кошек-самок компактные глыбки хроматина, которые но имени одного из исследователей получили в дальнейшем название телец Барра. В 1961 г. М. Лайон, изучая действие генов Х-хромосомы у мышей Mus musculus, выдвинула гипотезу об отсутствии генетической активности у гетеропикнотических Х-хромосом, которые в различных соматических клетках мышей могут быть либо материнского, либо отцовского происхождения.
Согласно этой гипотезе, в случае гетерозиготности по генам, сцепленным с полом, самки должны иметь мозаичный фенотип, поскольку содержат два типа клеток: одни - с мутантным, другие - с нормальным геном. Правильность этого предположения для Х-хромосом других млекопитающих, и в частности, человека была блестяще продемонстрирована на целом ряде примеров практически сразу после выдвижения гипотезы . Так в эритроцитах женщин, гетерозиготных по гену недостаточности глюкозо-6-фосфат-дегидрогеназы было обнаружено два типа клеток — с активным и неактивным ферментом. Подобный мозаичный фенотип был установлен по гену «недержания пигмента», по гену дефектной зубной эмали, а также по многим другим генам, локализованным в Х-хромосоме и отвечающим за Х-сцеплен-ные наследственные заболевания.
Многочисленые цитологические и биохимические исследования последующих лет подтвердили гипотезу М. Лайон и расширили описание феноменологии процесса, получившего название лайонизации Х-хромосомы самок. Эффект лайонизации был продемонстрирован с помощью разных методов. В частности, при цветовой слепоте обнаружили пятна дефектного цветовосприятия, используя для освещения сетчатки узкий пучок попеременно то красного, то зеленого света.
Современная теория инактивации Х-хромосомы. Инактивация одной из родительских Х-хромосом происходит в раннем эмбриональном развитии во всех соматических клетках самок млекопитающих. Она осуществляется путем гетерохроматизации: инактивироваиные Х-хромосомы, превращаются в плотно конденсированные тельца и втаком виде располагаются обычно по периферии ядра. Они образуют в соматических клетках гетеропикнотичные тельца Барра (половой хроматин). Ферменты транскрипции не могут работать на конденсированном хроматине, следовательно, дозы функционирующих генов становятся одинаковыми как у самцов, так и у самок. В случае полисомии по Х-хромосоме инактивируются все, кроме одной хромосомы, поэтому число телец Барра можно вsчислить по формуле n-1, где n - число Х-хромосом.
Неактивное состояние одной из Х-хромосом, установившись однажды в раннем эмбриогенезе, клонально передается дочерним соматическим клеткам во всех последующих клеточных поколениях. Предполагают, что роль механизма, закрепляющего инактивацию, играет метилирование цитозинов в молекуле ДНК.
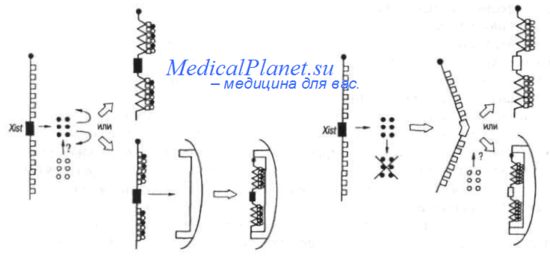
Инактивация в соматических клетках носит, как правило, случайный, характер, т.е. равновероятный для Х-хромосом материнского и отцовского происхождения. Однако есть немногочисленные примеры неслучайной инактивации одной из родительских Х-хромосом. В первую очередь к ним относятся аллели локуса Хсе у мышей -возможного кандидата на роль центра инактивации Х-хромосомы у этих млекопитающих. Преимущественная инактивация отцовской хромосомы выявлена в соматических тканях представителей отрядов яйцекладущих и сумчатых. В оогониях самок млекопитающих незадолго до вступления в профазу I мейоза неактивная Х-хромосома подвергается реактивации, что свидетельствует об обратимом характере инактивации в клетках этого типа. У самцов мыши единственная Х-хромосома в раннем сперматогенезе инактивирована и реактивируется на его конечных этапах. Цикл инактивации-реактивации Х-хромосомы у мыши представлен на рисунке, Он демонстрирует установленную экспериментально преимущественную экспрессию гена Xist на отцовской Х-хромосоме.
В настоящее время считается доказанным, что инактивация начинается в единственном центре Х1С (от англ. X-chromosome inactivation center), а затем прогрессивно распространяется вдоль всей длины хромосомы по закону «все или ничего». Все гены одной хромосомы инактивируются, а гены другой — остаются активными. Однако на Х-хромосоме человека в трех отдельных районах локализованы 8 генов, не повергающихся инактивации, причем один из них активен исключительно на инактивированной Х-хромосоме, а семь - на обеих.
На картах митотических хромосом человека и мыши локализованы центры инактивации. Предполагается, что Х-хромосома остается активной до тех лор, пока не получит сигнал инактивации из такого центра. В районе ДНК протяженностью 50 т.п.н., который он занимает, находятся 4 компонента: Хсе, Xist, Tsix и DXPas34, играющие важную роль в инактивации Х-хромосомы. Оказалось, что ген Хсе (от англ. X-chromosome-controlling element) мыши, представленный тремя аллелями, контролирует выбор для инактивации одной из родительских Х-хромосом.
Ген Xist (отангл. X-inactive-specific transcript) кодирует нетранслируемую РНК, необходимую для инактивации Х-хромосомы, Его экспрессия обнаруживается в тех клетках, где Х-хромосома неактивна, но отсутствует у самцов и особей ХО. У человека полная последовательность гена XIST, имеющего несколько сайтов начала транскрипции, составляет более полутора десятков т.п.н. и включает 8 экзонов.
Tsix представляет собой РНК, контролирующую экспрессию гена Xist. Эта. РНК транскрибируется с параллельной (антисмысловой) цепи ДНК гена Хist. Активация Tsix происходит под влиянием локуса DXPas34, содержащего 34-членный повтор саттелитной ДНК.
Существует несколько моделей инактивации Х-хромосомы с участием гена Xist. Часть из них предполагает существование двух факторов, один из которых (продуцируемый кратковременно и в очень ограниченном количестве) блокирует инактивацию, а другой запускает ее. Как один из вариантов рассматривается возможность инициации транскрипции гена ХШ в результате произошедшего связывания гипотетического сигнала с центром инактивации. Предполагается, что дальше Xist может инициировать гетерохроматизацию локализованных вблизи него участков Х-хромосомы. Согласно другой гипотезе что ген Xist у мышей представляет собой центр инактивации, ответственный за все ее этапы, начиная с выбора экспрессируемого аллеля и заканчивая распространением и поддержанием инактивации. Две модели, базирующиеся на этой возможности, представлены на рисунке.
Инактивация Х-хромосомы в соматических клетках у плацентарных млекопитающих чрезвычайно стабильна. Напротив, инактивация у сумчатых млекопитающих весьма лабильна: реактивацию Х-хромосомы можно обнаружить как in vivo, так и в клеточных культурах разных тканей. Таким образом, наблюдается межвидовая вариация стабильности инактивированного состояния Х-хромосомы и внутривидовая — среди разных Х-сцепленных генов.
Установлено наличие небольших групп генов (в псевдоаутосомных районах), остающихся активными в инактивированной Х-хромосоме и имеющих гомологов в Y-хромосоме. Кроме них, в коротком плече, и в дистальном отделе длинного плеча X-хромосомы есть гены, сохраняющие активность в инактивированной Х-хромосоме. У мыши соответствующие гены в норме инактивируются. Разница в составе инакти-вируемьгх генов у человека и мыши может объяснить разные последствия появления особей с аномальным числом Х-хромосом у этих видов. Так, у человека большинство индивидов женского пола с кариотипом ХО погибает на эмбриональной стадии развития, а выжившие имеют заметные фенотипические аномалии, тогда как самки ХО у мыши внешне совершенно нормальные. Наличие аномалий у женщин ХО может быть связано с недостаточным количеством продукта генов, исключенных из инактивации, вследствие отсутствия второй Х-хромосомы. Напротив, у индивидов с дополнительной Х-хромосомой врожденные пороки развития могут быть результатом избыточной дозы этих генов.
- Читать далее "Регуляция генной активности на уровне репликации. Трансляционная и посттрансляционная регуляция генной активности."
Оглавление темы "Генные и хромосомные мутации.":1. Регуляция транскрипции у прокариот. Негативная и позитивная регуляция генной активности.
2. Специфическая регуляция генной активности. Методы регуляции генной активности.
3. Неспецифическая регуляция генной активности. Компенсация дозы генов у дрозофилы.
4. Компенсация дозы генов у млекопитающих. Современная теория инактивации Х-хромосомы.
5. Регуляция генной активности на уровне репликации. Трансляционная и посттрансляционная регуляция генной активности.
6. Мутации. Теоритические основы мутационной изменчивости.
7. Геномные мутации. Гаплоидия. Полиплоидия.
8. Анеуплоидия. Нуллисомия. Моносомия. Полисемия.
9. Хромосомные мутации. Делеции. Дупликации.
10. Инверсии хромосом. Транслокации хромосом.