
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Элонгация цепей ДНК. Элонгация как этап синтеза ДНК.
Синтез новых цепей ДНК осуществляется группой ферментов с общим названием ДНК-полимеразы. ДНК-полимеразы многих организмов выделены и хорошо охарактеризованы. Все полимеразы способны образовывать ковалентную связь между 3'-концом цепи и 5'-концом свободного нуклеотидтрифосфата. Следовательно, новая цепь растет в направлении от 5'-конца к 3'-концу вдоль матричной цепи, имеющей противоположное направление 3' -» 5'. Реакцию присоединения нуклеотида эти ферменты осуществляют только в присутствии одноцепочечной матрицы и короткого двухцепочечного участка со свободным 3'-концом — праймера. Первый дезоксирибонуклеотид присоединяется к 3'-концу РНК-праймера.
Затем ДНК-полимераза один за другим присоединяет нуклеотиды, строя, таким образом, цепочку ДНК. Поскольку все полимеразы способны строить цепь только в одном направлении, на 3'-5' родительской цепи синтез новой цепи будет идти непрерывно, эту цепь принято называть лидирующей. Лидирующая цепь растет в направлении движения вилки, для ее элонгации необходим лишь один акт инициации. На другой цепи синтез осуществляется короткими фрагментами, которые называют фрагментами Оказакн по имени исследователя, впервые их обнаружившего. Эта цепь называется запаздывающей, или отстающей. Длина фрагментов Оказаки составляет 1000—2 000 п.н. у прокариот и 100—200 п.н. у эукариот. Отстающая цепь растет в направлении противоположном движению вилки, при ее синтезе необходимо множество актов инициации.
Фрагменты Оказаки объединяются в непрерывную цепь ДНК после удаления РНК-праймеров и застраивания освободившихся участков дезоксирибонуклеотида ми. Реакцию сшивания фрагментов осуществляют ферменты ДНК-лигазы, которые катализируют образование ковалентной связи между 3'-ОН одного фрагмента и 5'-фосфатом следующего.
Общая схема процессов, происходящих при элонгации цепей, одинакова у про- и эукариот.
У кишечной палочки в репликации ДНК участвуют два фермента: ДНК-полимераза I и ДНК-полимераза III. Ключевым ферментом репликации является ДНК-полимераза III. Полный комплекс ДНК-полимеразы III, способный осуществлять реакцию полимеризации, включает, по крайней мере, 20 разных белков, но каталитическую функцию выполняют 3 субъединицы - а (альфа), обладающая по-лимеразной активностью, е (эпсилон), обладающая 3' —> 5'экзонуклеазной активностью, и тета, функция которой пока неясна. Функции других субъединиц тоже неизвестны. Одна из них — В - уменьшает вероятность отделения фермента от матрицы до завершения процесса копирования. В каждой вилке репликации находится две молекулы холофермента, а в клетке -10-20 молекул. Он синтезирует ДН К лидирующей и отстающей цепей,
ДНК-полимераза I участвует в синтезе отстающей цепи. Этот фермент состоит из одной полипептидной цепи и имеет 3 ферментативные активности. Благодаря 5' —> З'-экзонуклеазной активности он удаляет РН К-праймер, отщепляя рибонуклеотиды с 5'-концов фрагментов Оказаки. Его полимеразная активность обеспечивает наращивание цепи ДНК предыдущего фрагмента. 3' -» 5'-экзонуклеазная активность позволяет контролировать правильность присоединения каждого нуклеотида и удалять ошибочно вставленные неспаренные нуклеотиды с растущего конца цепи.
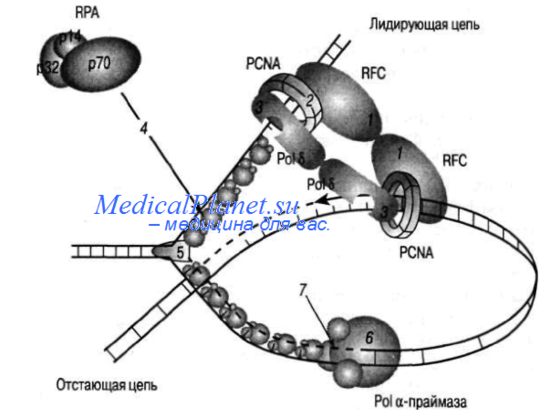
В репликативном синтезе ядерной ДНК эукариот участвуют два фермента ДНК-полимеразы. Наращивание лидирующей и отстающей цепей обеспечивает сложный белковый комплекс, в составе которого самое важное значение имеет 5-ДНК-полимераза, репликационный фактор RFC (от англ. replication factor С) и белок PCNA (от англ. proliferating cell nuclear antigen). Репликационный фактор RFC состоит из пяти субъединиц различной молекулярной массы (140/145,40, 38, 37 и 36.5 кДа). Этот белок связывается с 3'-концом только что синтезированного праймера и блокируетего наращивание, выполняемое а-ДН К-полимеразой) при длине примерно 30 нуклеотидов. На этой стадии RFC способствует связыванию ДНК с белком PCNА, который, по-видимому, играет рол ь связующего звена между полимеразами и другими белковыми факторами комплекса.
В результате этих процессов ДНК-полимераза вытесняется с 3'-конца растущей цепи. Дальнейший синтез продолжается другими ферментами - 6- и е-полимеразами. 5-ДНК-полимераза человека - мультимерный белок, состоящий из четырех субъединиц 125, 50, 68 и 12 кДа. Тройной комплекс RFC-PCNA-5-ДНК-полимераза обеспечивает элонгацию обеих цепей ДНК. Кроме репликации 5-ДН К палимераза участвует и в репаративных синтезах ДНК.
Удаление праймеров и сшивание фрагментов Оказаки происходит с участием 5-ДНК-полимеразы, РНКазы Н, ДНК-лигазы иф лэпэндонуклеазы. РНКаза Н катализирует расщепление РНК в гибридных РНК-ДНК-молекулах, флэп-эндонуклеаза отщепляет так называемые «свисающие 5'- концы», которые образуются в результате того, что 5-ДНК-полимераза обеспечивает синтез с вытеснением цепи.
Синтез митохондриальной ДНК осуществляету-ДНК-полимераза. Но механизм репликации ДНК в митохондриях несколько отличается от репликации хромосомной ДНК прокариот и эукариот. Наиболее важная особенность репликации митохондриальной ДНК - наличие сайтов инициации, специфичных для каждой из комплементарных цепей ДНК. Ген POLG, кодирующий у-ДНК-полимеразу у человека, локализован в 15q25. Праймер синтезируется митохондриальной ДНК-зависимой РНК-полимеразоЙ, отвечающей как за экспрессию митохондриальных генов, так и за создание РНК-праймера для инициации репликации митохондриального генома. У человека этот белок кодирует ген POLRMT, локализованный в 19р13.3. Продукт этого гена больше похож на РНК-полимеразы фагов и митохондриальные полимеразы низших эукариот.
Другие многочисленные ДНК-полимеразы эукариот обеспечивают синтез ДНК, необходимый для репарации повреждений ДНК и рекомбинации.
- Читать далее "Репликация ДНК. Характеристика параметров репликации."
Оглавление темы "Репликация ДНК - виды и механизмы.":1. Элонгация цепей ДНК. Элонгация как этап синтеза ДНК.
2. Репликация ДНК. Характеристика параметров репликации.
3. Двунаправленность репликации ДНК. Полирепликонность и асинхронность репликации.
4. Восстановление или репарация ДНК. Нарушение первичной структуры ДНК.
5. Виды репарации ДНК. Фотореактивация.
6. Репарация ДНК за счет ДНК-полимераз. Эксцизионная репарация ДНК.
7. Исправление ошибок спаривания ДНК. Мисмэтч-репарация. Рекомбинационная репарация.
8. SOS репарация ДНК. Характеристика и механизмы SOS репарации ДНК.
9. Репарация ДНК и наследственные болезни. Пигментная ксеродерма.
10. Синдром Коккейна. Трихотиодистрофия. Генетические основы синдрома Коккейна и трихотиодистрофии.