
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Сравнение карт. Неравный кроссинговер. Соматический кроссинговер.
В целом, сравнение генетических и цитологических карт хромосом показывает их соответствие: чем больший процент кроссинговера разделяет пару генов, тем больше и физическое расстояние между ними. Порядок расположения генов на этих картах совпадает, однако расстояния между генами могут заметно различаться. Это связано с тем, что кроссинговер в прицентромерных и теломерных районах у дрозофилы затруднен, поэтому и расстояния между генами на генетической карте в этих районах занижены.
Кроме того, частота кроссинговера зависит от ряда факторов: температуры, возраста особей, их пола, жизнеспособности, а также присутствия генов, влияющих на частоту кроссинговера, и структуры хромосом. Вследствие этого определение частоты кроссинговера между одними и теми же генами у разных авторов может не совпадать и, кроме того, не соответствовать расстоянию на цитологической карте хромосом.
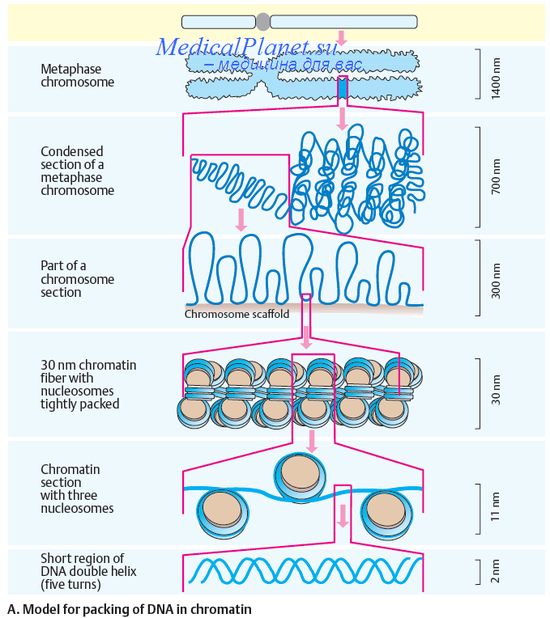
В настоящее время цитологические карты хромосом построены у различных организмов, в том числе и у человека. Это стало возможным благодаря использованию специальных методов дифференциального окрашивания хромосом с помощью флуоресцентных красителей или по Гимза. Эти методы выявляют на каждой хромосоме окрашенные и неокрашенные сегменты. При обозначении местоположения гена сначала указывается номер хромосомы, в которой он локализован, затем символ плеча (р - короткое и q — длинное), номер района и номер сегмента в пределах этого района.
Например, запись 1q2l означает, что речь идет о хромосоме 1, длинном плече, районе - 2, сегменте - j. Такая номенклатура сегментов позволяет описать цитологическую локализацию гена. Например, ген миодистрофии Дюшенна локализован в коротком плече Х-хромосомы Хр21.
Развитие молекулярной биологии привело к появлению новых современных методов картирования генов. Один из них - ДНК-гибридизация in situ. Для локализации какого-либо гена получают информационную РН К, транскрибированную с этого гена, а затем с помощью обратной транскриптазы ее ДНК-копию (кДНК - комплементарная ДНК). После этого проводят гибридизацию с денатурированной хромосомной ДНК и по месту гибридизации определяют, в каком участке хромосомы локализован данный ген.
Неравный кроссинговер.
Обычно обмен участками между хроматидами гомологичных хромосом осуществляется в строго идентичных, тождественных точках. Благодаря этому при перекресте происходит обмен равными участками хромосом.
В очень редких случаях наблюдаются разрывы в несимметричных точках, и хроматиды обмениваются неравными участками. Такое явление называют неравным кроссинговером. Вследствие неравного кроссинговера участок одной из гомологичных хромосом может удвоиться или утроиться, а в противоположной хромосоме образуется его нехватка. Например, изучая мутацию Ваr (полосковидные глаза) у дрозофилы, Стертевантв 1925 г. обнаружил, что фенотип «полосковидные глаза» у таких мух обусловлен удвоением участка 16А в первой хромосоме.
Но среди мух, несущих мутацию Ваr, иногда выявлялись особи с еще более узкими глазами и мухи с нормальными глазами. Оказалось, что хромосомы особей ультра-Ваr имеют утроенный участок 16А, у мух дикого типа хромосомы нормальные. По мнению Стертеванта, такие хромосомы могли образоваться в результате неравного кроссинговера по участку 16А у гомозиготных особей В/В.
Соматический кроссинговер.
Известно, что гомологичные хромосомы в профазе митоза обычно не вступают в си -наптическую связь, а располагаются отдельно и независимо друг от друга. В то же время описаны случаи, когда наблюдается митотическая конъюгация гомологичных хромосом и рекомбинация между несестринскими хроматидами.
Доказательство существования митотического кроссинговера было получено при наблюдении мозаичных пятен у мух с генотипом (у sn+/y+ sn). Пятна образуются тогда, когда рядом расположены два клона клеток, фенотипически отличающихся друг от друга и от клеток остальных тканей данной особи. Гены у (yellow — желтое тело) и sn (singed - «опаленные» щетинки) находятся в Х-хромосоме. Самка у sn+/y+ sn гетерозиготна по генам у и sn, и поэтому в отсутствие митотического кроссинговера ее фенотип будет нормальным.
Однако если на стадии четырех хроматид между хроматидами гомологов (несестринскими хроматидами) произойдет кроссинговер, причем место обмена будет находиться между геном sn и центромерой, то после завершения деления могут образоваться клетки с генотипами у sn+/y sn+ и у+ sn/y+ sn. Для этого необходимо, чтобы после кроссинговера хроматиды каждого из гомологов отошли к одному полюсу клетки: y+sn- к одному полюсу, aysn* - к другому. Потомки дочерних клеток, размножившись в имагинальных дисках на стадии личинки, образуют мозаичные пятна.
В этом случае на сером теле мухи с нормальными щетинками появятся два пятпа, одно из которых будет желтого цвета с нормальными щетинками, а другое -серого цвета с «опаленными» щетинками.
- Читать далее "Частота кроссинговера. Факторы влияющие на кроссинговер."
Оглавление темы "Генотип и кроссинговер.":1. Генотип и фенотип. История изучения генотипа и фенотипа.
2. Вариация наследственных признаков. Фенотип и развитие организма.
3. Гетерогенные группы. Генокопии и фенокопии. Неоморфизм. Гипоморфизм. Антиморфизм.
4. Пенетрантностъ признака. Экспрессивность признака.
5. Неустойчивая и условная доминантность в генетике. Множественные аллели признака.
6. Сцепленное наследование. Полное сцепление. Неполное сцепление. Кроссинговер.
7. Расстояние между генами. Двойной кроссинговер. Интерференция генов.
8. Генетические карты. Картирование генов.
9. Сравнение карт. Неравный кроссинговер. Соматический кроссинговер.
10. Частота кроссинговера. Факторы влияющие на кроссинговер.