
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Индукция и органогенез мочеточника и почки.
Ген, кодирующий корецептор для Ret — Gfral (производный глиальной линии клеток нейротрофический рецептор фактора), также экспрессируется в зачатке мочеточника, и его инактивация у мышей ведет к появлению фенотипа, подобного наблюдаемому у мышей с «угнетенными» генами Ret и Gdnf, что подтверждает роль Gfral в передаче сигналов Gdnf. Недавние исследования показали, что две изоформы Ret (Ret9 и Ret5l) имеют различное действие in vivo: моноизоформные Ret51 мыши, у которых нет Ret9, характеризуются гиподисплазией почки и недостаточностью кишечных ганглиев в толстом кишечнике.
Возможно, Rel/Gfral/Gdnf-система координирует рост мочеточника с помощью других секретируемых факторов, к которым относится, в частности, белок Втр4, стимулирующий морфогенез костной ткани. Ген Втр4 экспрессируется в клетках мезенхимы, которые окружают вол ьфов проток и ствол мочеточника в месте, удобном для регуляции роста данного зачатка. Эмбрионы мыши с генотипом Втр4+/- имеют целый ряд аномалий развития, включая дефекты почек, вызванные нарушением регуляции развития зачатка мочеточника, причем эти аномалии подобны врожденным порокам развития почки и мочевого тракта у человека. В клеточной культуре Втр4 регулирует гены, которые экс-прессируются и в мочеточниковом зачатке, и в мезенхиме, включая Gdnf.
Белки поверхности клетки, такие как протеогликаны (PGs), также вовлечены в регуляцию роста и разветвления мочеточника в процессе развития почки. PGs клеточной поверхности принимают участие в передаче сигналов фактора роста фибробластов (Fgfs) и входят в семейство факторов Wnt. При удалении или изменении в почке цепей PGs in vitro с помощью химических реагентов, модифицирующих синтез, зачаток мочеточника приостанавливает свой рост и ветвление. Эти преобразования могут также вызывать полную реверсию действия Втр4: от подавления роста зачатка мочеточника к стимулированию его ветвления, что указывает, на возможность для PGs контролировать специфический тип ответа на факторы роста. Вопрос об участии специфических PGs в развитии почки вышел на первый план после экспериментов по выключению (нокаутированию) у мышей гена одного из протеогликанов (Gpc3)t который также видоизменен у людей с синдромом Симпсона-Голаби~Бемеля и почечной дисплазией.
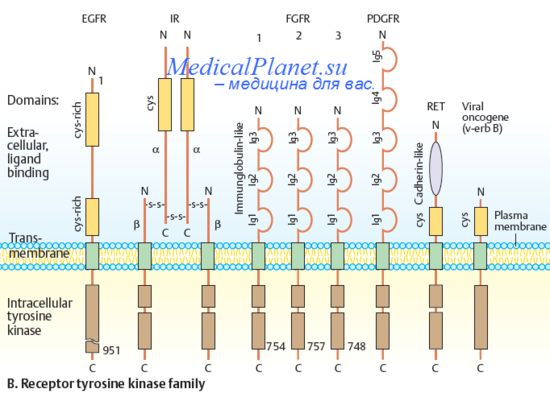
Системе сигнализации Gdnf/Ret принадлежит решающая роль в индукции мочеточников и их ветвлении, но она может модулироваться другими факторами, способствуя генерации специфического паттерна в древе ветвления мочеточников. Начиная с пионерских работ К. Гробстайна, установлено, что почечные канальцы индуцированы тканевыми взаимодействиями. Формирование трубки в мезенхиме вызвается in vivo зачатком мочеточника, a in vitro - различными индукторами, например, спинным мозгом. Классическое представление об индукторах состояло в том, что мезенхимные клетки, которые входят в контакте зачатком мочеточника и его сигналом, продолжают формирование почечных канальцев. За последние годы, однако, и в эпителии мочеточников, и в мезенхиме были идентифицированы гены, необходимые для нефрогенеза, играющие решающую роль и для сигналов, генерируемых зачатком мочеточника, и для сигналов — производных мезенхимы.
Эти гены также определяют нисходящие сигналы в почечной мезенхиме, управляющие нефрогенезом.
В экспериментах по инактивация гена Етх2 у мышей был получен материал для модели, в которой сигнал зачатка мочеточника участвует в индукции канальца in vivo. Как гомеобокс-содержащий фактор транскрипции, который экспрессируется первоначально в зачатке мочеточника, Етх2 может также регулировать экспрессию других генов, вносящих вклад в нарушение тубулогенеза при отсутствии Етх2. У Emx2-нокаутных мышей, зачаток мочеточника внедряется в мезенхиму метанефроса, однако индукция канальцев отсутствует и, следовательно, развития почки не происходит. Причиной является то, что, хотя некоторые мезенхимные и эпителиальные маркерные гены раннего почечного развития, такие как Wtl, Gdnf и Ret, экспрессированы у Emx2-мутантов, в клетках их мезенхимы отсутствует экспрессия Wnt4 -важного первоначального сигнала для тубулогенеза.
Wnt — большое семейство секретируемых сигналов, которые регулируют ключевые морфогенетические шаги входе эмбриогенеза. В 1994 г. было установлено, что Wnt1 может дублировать влияние спинного мозга (в пробах почечной культуры органа), играя роль индуктора тубулогенеза. С тех пор Wnts отнесены к тубуло-индуктивным сигналам. Однако в норме Wnt1 не экспрессируется в эмбриональной почке. Данный факт указывает, что он дублирует сигнализацию другого члена этого семейства, который экспрессируется в почечной мезенхиме или зачатке мочеточника. Действительно, в уплотненной мезенхиме и претубулярных агрегатах экспрессируется Wnt4, который, вероятно, и является тем мезенхимным сигналом, который участвует в эпителио-мезенхимном переходе. Интересно, что клетки, экспрессирующие ряд Wnt, были в состоянии стимулировать тубулогенез в почечной мезенхиме in vitro. Клетки, экспрессируюшие Wnt4, могли также преодолеть дефект тубулогенеза в мезенхиме Wnt4-мутантов.
Эти исследования указывают на важность тубуло-стимулируюшей роли Wnt4 и свидетельствуют, что классическая, стимулирующая почку, фильтрующая проба, при которой почечный тубулогенез вызван совместным культивированием эмбриональной почки и части спинного мозга, может действовать благодаря тому, что спинной мозг производит сигнал Wnt. Другой Wnt - Wnt6, экспрессируется в развивающейся почке, на концах зачатка мочеточника, что делает его возможным индуктором восходящей Wnt4-регуляции.
Важными регуляторами развития являются и ростовые факторы Вmр, принадлежащие Tgf-суперсемейству выделяемых сигналов. Хотя некоторые Втр экспрессируются в почке, первое генетическое доказательство того, что они функционируют в течение нефрогенеза, было получено путем нокаутирования Втр7 у мышей. Почки у них сначала формируются как обычно, происходит и ветвление зачатка мочеточника, но, начиная с 14.5 дня развития, нефрогенез прекращается. Экспрессия Рах2, Wtl и Wnt4 снижена в мезенхиме метанефроса у мутантов, и это может объяснять усиленный апоптоз, который замечен в почках Втр 7-нокаутных эмбрионов. Таким образом, Втр7 может функционировать в качестве сигнала выживания, который предотвращает подверженность мезенхимных клеток апоптозу.
Показано, что дополнительный Вmр7 в культуре предотвращает подверженность апоптозу клеток дикого типа в мезенхиме метанефроса, но такие мезенхимные клетки не способны формировать канальцы. В соединении с Fgf2, однако, Вmр7 может стимулировать рост клеток стромы и поддерживать способность мезенхимы метанефроса мезенхимы подвергаться нефрогенезу in vitro. На основе этих результатов было высказано предположение, что нефрогенная мезенхима служит источником сигналов, которые способствуют быстрому увеличению стромальных клеток-предшественников. Эти клетки в свою очередь, выделяют еще не идентифицированные анти-дифференциационные сигналы и сигналы выживания, контролирующие интенсивность нефрогенеза.
В настоящее время известно, что и эпителиальные, и мезенхимные гены регулируют индуктивные взаимодействия в почке, сигнализация Wnt идентифицирована как критический индуктор тубулогенеза. Вероятно, что активность Wnt модулируется другими проводящими путями, такими как Notch и Вmр, что остается предметом дальнейшего изучения.
В дополнение к зачатку мочеточника и нефрогенным центрам сигнализации мезенхимы важным источником сигнализации в почечном органогенезе является строма. Первое свидетельство значения стромы было получено при изучении экспрессирующегося в ней нокаутированного Foxd1-rem. Хотя его инактивация ведет к дефектам в системе собирающего протока и нефронов, он не экспрессируется в клетках названных тканей. Это позволило предположить, что Foxd1 может участвовать в почечном развитии, регулируя экспрессию сигналов, продуцируемых клетками стромы. Данные сигналы способствуют тубулогенезу и развитию зачатка мочеточника. Сигнализация клеток стромы поддерживает экспрессию гена Ret, которая, в свою очередь, подключает сигнализацию, регулирующую дифференцировку мезенхимных клеток. Эти данные указывают, что между стромой, зачатком мочеточника и почечной мезенхимой может существовать сигнальное кольцо, которое коордипирует почечное развитие.
Стромальная сигнализация может также влиять на нефрогенную мезенхиму или на зачаток мочеточника, а сигнализация от нефрогенной зоны может, в свою очередь, регулировать компартмент стромы. Анализ мутантных фенотипов дает обширную информацию о природе клеток и взаимодействии тканей, которые управляют ранним развитием почки.
Что касается молекулярной природы индукторов у других модельных организмов, то гомологи трех из числа названных в таблице генов играют важную роль в индукции образования мезодермы уже упоминавшейся африканской шпорцевой лягушки Xenopus laevis. Это гены семейств BMP, FGF и Wnt. Так, в результате экспрессии гена ВМР4 индуцируется лорсо-вентральная организация мезодермы, ген FGF вовлечен в индукцию вентро-латеральной мезодермы, а гены семейства wnt Xenopus (Xwnt-1 и Xwnt-S) играют существенную роль в индукции формирования полной дорсальной оси зародыша Xenopus. Экспрессия Frizzled-связанного белка (sFrp), который противодействует эффектам индукции Xwnt8 у лягушек и, очевидно, предотвращает активацию рецептора, также коррелирует с экспрессией Wmt4 в почке мыши.
Кроме того, два гомолога относятся к нейроиндуцируюшим факторам. Это - ген Xotch у шпорцевой лягушки, продукт которого локализуется в дорсальной эктодерме нейрулы. У дрозофилы его гомолог — ген Notch, участвует в выборе клетками нейроэктодермы нейрального или эпидермального пути развития. Другой ген Wnt-1 экспрессируется в области среднего и заднего мозга и участвует в индукции клеточной пролиферации,
Изучение почки позволило уточнить механизмы специализации эпителиальных клеток и взаимных индуктивных взаимодействий, которые существуют между различными типами клеток и тканей, а также вклад, который эти взаимодействия вносят на ранних этапах развития. Некоторые «почечные» мутанты мышей, оказались также полезными моделями для изучения наследственных болезней почек и расстройств у человека.
- Читать далее "Популяционная генетика. Что такое популяция?"
Оглавление темы "Популяционная и молекулярная генетика.":1. Гены контролирующие индукцию. Индукция и органогенез.
2. Индукция и органогенез мочеточника и почки.
3. Популяционная генетика. Что такое популяция?
4. Закон Харди—Вайнберга. Условия закона Харди—Вайнберга.
5. Инбридинг. Панмиксия. Отсутствие панмиксии.
6. Дрейф генов. Мутационный процесс в популяции.
7. Миграции в популяции. Отбор в популяции.
8. Генетический полиморфизм в популяции. Генетический груз популяции.
9. Выделение ДНК. Химический синтез ДНК.
10. Полимеразная цепная реакция. Принципы ПЦР.