
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Регуляция силы сокращения миокарда. Влияние массажа сердца на его активность
Как известно, синхронная регистрация такого суммарного потенциала с мембранным потенциалом одной из клеток миокарда, расположенных в этой же зоне (мембранный потенциал клеток регистрировался микроэлектродом), свидетельствует о значительном совпадении фазы и формы обоих потенциалов. Однако амплитуда потенциала действия одной клетки является значительно большей, чем амплитуда монофазного потенциала, отражающего суммарную активность многих клеток (Гоффман, Крейнфилд, 1964). Это различие связано с тем, что между клетками, генерирующими потенциал, и электродом находится слой ткани, обладающий сопротивлением и способностью шунтировать потенциалы отдельных клеток. Явление это аналогично тому, которое возникает при регистрации потенциалов действия целого «нервного ствола и отдельных его волокон.
Известно, что при сильных раздражениях нервного ствола амплитуда монофазных потенциалов возрастает, что расценивают как следствие вовлечения в возбуждение большего количества нервных волокон. Изменение величины суммарного потенциала позволяет в некоторой мере судить об изменениях количества нервных волокон, синхронно охваченных процессом возбуждения (Guyton, 1961).
Мы попытались использовать подобный прием для изучения вопроса о возможности изменения количества волокон миокарда, синхронно охватываемых процессом возбуждения. Указанным выше способом регистрировался монофазный потенциал поверхности левого желудочка. Одновременно регистрировалось давление в устье аорты, позволившее судить об изменениях силы сокращения миокарда левого желудочка. При этом выявлено, что воздействия, дающие положительный инотропный эффект, т. е. увеличение силы сокращения левого желудочка (в том числе и введение в сердце адреналина), увеличивают амплитуду монофазных потенциалов миокарда левого желудочка.
Известно, что возбуждение симпатической нервной системы, введение адреналина (и другие воздействия, дающие положительный инотропный эффект) не вызывают изменений величины потенциалов действия отдельных клеток миокарда желудочков (Гоффман, Крейнфилд, 1964). Следовательно, «наблюдавшееся нами увеличение амплитуды суммарного монофазного потенциала свидетельствует об увеличении количества клеток миокарда, синхронно охватываемых возбуждением при каждом сердечном цикле.
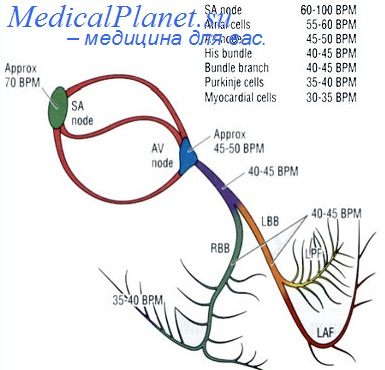
Можно думать поэтому, что сила сокращений миокарда может регулироваться путем градуального включения в функцию различного количества отдельных мышечных клеток. Такой градуальный принцип включения в деятельность отдельных функционалыных единиц соответствует основным чертам архитектуры физиологических функций (Д. Баркрофт, 1937) и обладает рядом преимуществ, обеспечивая более тонкую регуляцию силы сокращений миокарда в целом, а также возможность лучшего восстановления работоспособности отдельных мышечных клеток благодаря поочередному включению их в работу.
В пользу подобного предположения могут свидетельствовать наблюдения над характером восстановления сократительной активности миокарда при массаже сердца. На сердце кошки, изолированном по Лангендорфу, в некоторых опытах наступало резкое ослабление силы сокращений, несмотря на достаточный уровень перфузии коронарных сосудов артериальной кровью кошки-донора. При этом сохранялись обычный синусовый ритм и электрическая активность всех отделов сердца, однако сила сокращений миокарда была настолько слабой, что давление в баллончиках, находящихся в желудочках, при систолах повышалось лишь на несколько миллиметров ртутного столба. Легкий массаж сердца, а иногда даже простое прикосновение к нему в этих случаях приводило к восстановлению нормальной силы сокращений миокарда и подъему давления в правом и левом желудочках до 20—30 и 80—120 мм рт. ст. соответственно. В некоторых опытах также сильные удары возникали сразу в ответ на первые же прикосновения к сердцу. Ни о каком влиянии на коронарный кровоток здесь
не может быть и речи, так как на всем протяжении опыта перфузионный насос поддерживает постоянный и достаточно высокий уровень коронарного кровотока. Создается впечатление, что раздражение механорецепторов в этих случаях приводило к немедленному синхронному сокращению всех (или почти всех) клеток миокарда, что и вызывало мгновенное восстановление силы сокращений.
По-видимому, в этом и заключается одна из существенных причин благотворного действия массажа сердца на сократительную активность миокарда.
Сказанное подтверждается опытами с регистрацией момофазных потенциалов.
- Читать далее "Влияния на монофазные потенциалы. Структура нервной системы сердца"
Оглавление темы "Медиаторы сердечной нервной регуляции":1. Скорость проведения через атриовентрикулярный узел. Асинхронизм волокон миокарда
2. Альтернирующий пульс. Монофазные потенциалы миокарда
3. Регуляция силы сокращения миокарда. Влияние массажа сердца на его активность
4. Влияния на монофазные потенциалы. Структура нервной системы сердца
5. Передача возбуждения в сердце. Возбуждение и торможение нервной системы сердца
6. Медиаторы регуляции сердечной деятельности. Перфузионное давление сердца
7. Виды эфферентных нейронов сердца. Адренергические нейроны сердца
8. Внутрисердечные ганглии нервной системы. Тормозные мотонейроны нервной системы
9. Взаимодействие внутрисердечной нервной системы. Интракардиальные регуляторные механизмы
10. Взаимодействие регуляторных систем сердца. Фоновое раздражение сердца