
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Функциональная структура миокарда. Регуляция силы сокращения миокарда
В электрофизиологических опытах В. А. Шидловского и Н. С. Дауэ (1953) были выявлены факты, свидетельствующие, по-видимому, о том, что мышцу сердца нельзя считать синцитием в буквальном смысле этого слова. Такое же представление о структуре сердечной мышцы вытекает и из данных электронно-микроскопических исследований сердца, проведенных в последние годы.
Ruska (1965) показал, что сердечная мышца не является синцитием. Она состоит из отдельных мышечных клеток, размер которых намного меньше, нежели размер симпластов поперечно-полосатой мышцы. Мышечные клетки миокарда соединяются одна с другой с помощью вставочных дисков. Соседние участки мембран двух клеток в этих звеньях расположены очень близко друг к другу.
При увеличении в 100 000 раз можно обнаружить, что в некоторых щелях между двумя соседними мембранами практически нет никакого межклеточного пространства и они плотно прилегают одна к другой. В других щелях ширина этого пространства достигает даже 100 А. По-видимому, подобные типы контактов представляют собой синапсы с электрической передачей возбуждения, т. е. то, что принято называть эфапсами (Экллс, 1966).
Так как ширина щели в различных вставочных дисках колеблется в сравнительно широких пределах, можно думать, что и величина электрического сопротивления этих образований может быть различной. По-видимому, именно поэтому некоторые авторы полагают, что сопротивление этих вставочных дисков является большим, «ежели обычное электрическое сопротивление мембраны, что затрудняет переход возбуждения с одной мышечной клетки на другую (Гоффман и Крейнфилд, 1962; Sperelacis, 1963; Tarr, Sperelacis, 1964; Ruska, 1965).
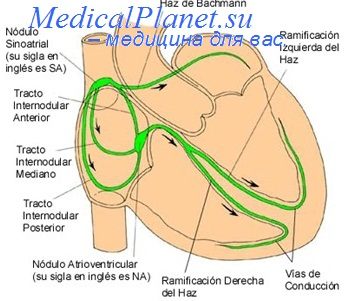
В то же время другие авторы (Вагг, 1965; Crill, Woodbury, 1962 a. oth.) полагают, что сопротивление вставочных дисков очень низкое, поэтому диски не затрудняют переход возбуждения с одной клетки миокарда на другую. В «последнем случае можно думать, что миокард в функциональном отношении представляет собой синцитий.
Поскольку ширина щели в различных дисках бывает разной, можно предположить, что одни вставочные диски обладают высоким сопротивлением, а другие — низким. В связи с этим, возможно, что клетки миокарда могут быть объединены в несколько групп «синцитиальных» структур. Maekawa, Kawamura и Hayashi (1965) обнаружили, что волокна Пуркинье также имеют клеточное строение и межклеточные соединения в этих волокнах, равно как и соединения между клетками волокон и клетками сократительного миокарда, по своей структуре являются такими же, как и вставочные диски между отдельными мышечными клетками миокарда.
Не исключена возможность, что величина сопротивления отдельных дисков может регулироваться каким-то механизмом, в связи с чем в процесс возбуждения может вовлекаться большее или меньшее количество клеток миокарда. В этом случае логично допустить, что сила сокращения миокарда (помимо факторов, описанных ранее) может регулироваться и посредством включения в сокращение большего или меньшего количества мышечных клеток. Еще Гаокел полагал, что возникновение перемежающегося пульса (pulsus alternans) зависит от включения в деятельность лишь части мышечных волокон желудочка при каждом втором ударе его.
Частичная асистолия миокарда связана с нарушением проведения возбуждения, а также, по-видимому, с различной длительностью рефрактерной фазы в разных волокнах. На основании ряда серий экспериментальных исследований и сопоставления их с литературными данными Н. В. Данилов (1929) пришел к выводу, что перемежающийся пульс возникает вследствие неполного включения в процесс возбуждения всех волокон миокарда желудочка. Он полагает, что эти волокна могут функционировать поочередно, в целях экономии сил миокарда. Поочередные включения волокон связаны с функцией системы, проводящей возбуждение.
- Читать далее "Сокращения миокарда. Нервная регуляция активности миокарда"
Оглавление темы "Регуляция сократимости миокарда":1. Старлинговский эффект. Регуляция изгнания крови
2. Отрицательные инотропные реакции на сердце. Отрицательные инотропные влияния на сердце
3. Механорецепторы каротидного синуса. Значение интракардиальных рефлексов в поддержании гемодинамики
4. Нервные регуляторные механизмы сердца. Экстракардиальные влияния на сердце
5. Регуляция возбудимости миокарда. Влияние сердечной нервной системы на возбудимость
6. Вызов экстрасистол сердца. Инотропные реакции сердца
7. Регуляция диастолического расслабления миокарда. Регуляция коронарного кровообращения
8. Расширение коронарных сосудов. Рефлекторная регуляция коронарных сосудов
9. Функциональная структура миокарда. Регуляция силы сокращения миокарда
10. Сокращения миокарда. Нервная регуляция активности миокарда