
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Фосфатидилинозитол-3-киназа (PI3-киназа) зависимая передача сигнала
Фосфатидилинозитол-3-киназа (PI3-киназа) связывается с фосфорилированными остатками тирозина и связанным IRS1 (субстратом инсулинового рецептора 1) активированного фактора роста и рецепторов инсулина. РI3-киназа образует PI3,4,5Р3 (фосфатидилинозитол-3,4,5-трифосфат), который фиксирован к клеточной мембране. PI3,4,5Р3 связывается с PDK1 (фосфоинозитидзависимая киназа 1) и протеинкиназой В (PKB/Akt).
В последующем PDK1 фосфорилирует и таким образом активирует PKB/Akt.
PKB/Akt стимулирует некоторые транспортные процессы, в т. ч. переносчик глюкозы GLUT4. Он фосфорилирует и, соответственно, инактивирует антипролиферативный и проапоптотический фактор транскрипции семейства Forkhead FKHRL1 (FoxO1), тем самым способствуя клеточной пролиферации и препятствуя апоптозу. В последующем РКВ/Akt фосфорилирует и активирует MDM2, который ингибирует проапоптотический фактор транскрипции р53.
Далее PDK1 и PKB/Akt регулируют экспрессию генов через фактор транскрипции NFkB. NFkB связан с ингибирующим белком IкВ и удерживается за счет этого в цитозоле. IкВ фосфорилируется 1кВ-киназой (IKK), что приводит к его убиквитинированию и деградации. При отсутствии IkB NFkB перемещается в ядро и стимулирует экспрессию генов. NFkB усиливает также синтез белков внеклеточного матрикса и, следовательно, способствует развитию фиброза.
PKB/Akt фосфорилирует и тем самым активирует IKK, что приводит к активации NFkB. Далее IKK активируется TNF-а и IL-1.
PKB/Akt фосфорилирует Bad — белок, стимулирующий освобождение цитохрома С из митохондрий, и с помощью этого запускает апоптоз. Фосфорилированный Bad связан с белком 14-3-3 и поэтому защищен от взаимодействия с митохондриями. PKB/Akt фосфорилирует и вследствие этого инактивирует каспазуЭ — протеазу, которая также участвует в сигнальном каскаде, ведущем к апоптозу. Таким образом, PKB/Akt подавляет апоптоз.
PKB/Akt фосфорилирует и с помощью этого активирует NOS. NO может аналогичным образом ингибировать апоптоз. PKB/Akt активирует p47Phox и в результате стимулирует образование активных форм кислорода (АФК).
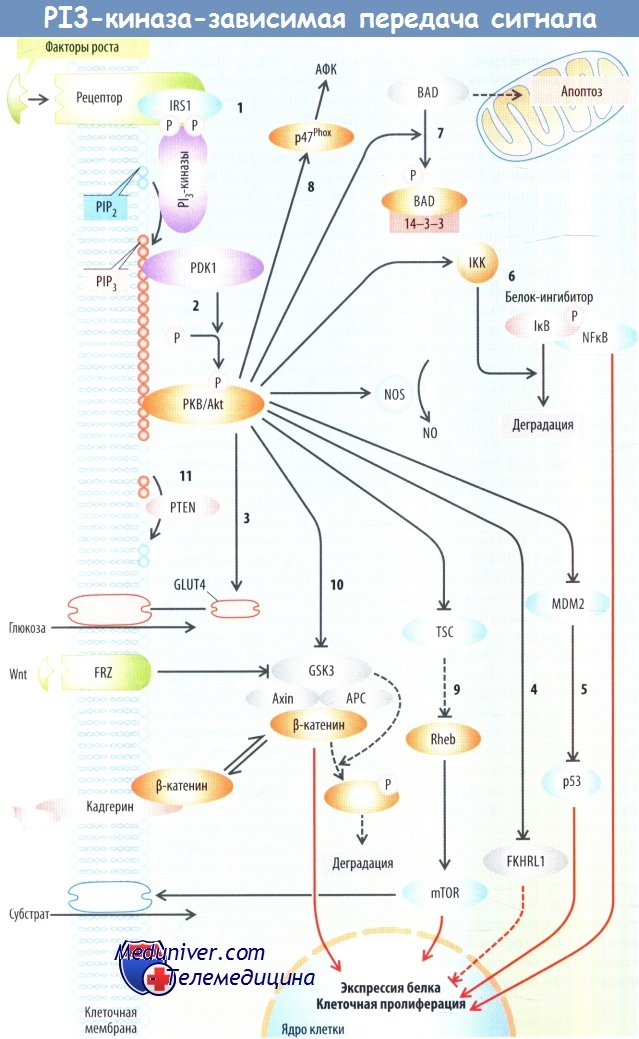
PKB/Akt фосфорилирует и за счет этого инактивирует туберин, который образует комплекс с гамартином (комплексом туберозного склероза — TSC). TSC инактивирует малый G-белок Rheb. Активированный Rheb стимулирует киназу mTOR (мишень рапамицина млекопитающих), белок, стимулирующий усвоение клеточного субстрата, белковый синтез и клеточную пролиферацию. Подавление туберина под действием PKB/Akt стимулирует mTOR.
Далее АМФ-активированная киназа (АМРК) стимулирует и тем самым ингибирует TSC. Истощение энергетических запасов клетки повышает концентрацию клеточного АМФ, а следовательно, активирует АМРК и ингибирует mTOR.
PKB/Akt фосфорилирует и с помощью этого инактивирует киназу 3 гликогенсинтазы (GSK3a и GSK3P). GSK3 в последующем ингибируется фактором роста Wnt, что происходит с участием рецепторов семейства Frizzled и белков семейства Disheveled. GSK3 соединяется с белковым комплексом, который состоит из Axin, белка Гиппеля—Линдау (vHL) и аденоматозного полипоза толстой кишки (АРС).
Этот комплекс связывается с многофункциональным белком β-катенином. GSK3 фосфорилирует β-катенин и этим запускает его деградацию, β-катенин может связываться с Е-кадгерином, который обеспечивает контакт между соседними клетками. Свободный β-катенин перемещается в ядро, взаимодействует с комплексом транскрипции TCF/Lef и благодаря этому стимулирует экспрессию нескольких генов, необходимых для клеточной пролиферации. Wnt и активированный PKB/Akt способствуют клеточной пролиферации отчасти за счет подавления GSK3 и последующей стимуляции β-катенинзависимой экспрессии генов.
PDK1 фосфорилирует и тем самым активирует киназу, индуцируемую сывороткой и глюкокортикоидами (SGK1). Экспрессия SGK1 стимулируется глюкокортикоидами, альдостероном, TGF-β, ишемией и гиперосмолярностью. SGK1 активирует ряд переносчиков, каналов и Na+/K+-АТФазу. Данная киназа имеет несколько таргетных белков, общих с PKB/Akt. После стимуляции их экспрессии она может играть ведущую роль в РI3К-зависимой передаче сигнала.
Фосфатаза PTEN дефосфорилирует PI3,4,5Р3 и таким образом завершает PI3,4,5Р3-зависимую передачу сигнала. Следовательно, PTEN ингибирует клеточную пролиферацию. Оксидативный стресс инактивирует PTEN и за счет этого увеличивает активность Akt/PKB и SGK.
- Рекомендуем ознакомиться со следующей статьей "Схема механизма некроза клетки"
Оглавление темы "Патофизиология в схемах":- Физиология роста клетки и ее адаптации
- Схема внутриклеточной передачи сигнала и его нарушений
- Фосфатидилинозитол-3-киназа (PI3-киназа) зависимая передача сигнала
- Схема механизма некроза клетки
- Схема механизма апоптоза клетки
- Схема образования опухолевых клеток - онкогенеза
- Схема влияний опухоли на организм - паранеопластических синдромов
- Схема регуляции старения организма человека и длительности его жизни
- Схема повышения температуры тела - лихорадки
- Схема перегревания организма (теплового удара) и злокачественной гипертермии