
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Механизмы отторжения трансплантатов органов
Отторжение представляет собой комплексный процесс, в котором играют роль как клеточный иммунитет, так и циркулирующие антитела. Вклад этих механизмов часто отражает гистологическая картина отторженного органа.
а) Реакции, опосредуемые Т-клетками. Важнейшая роль Т-клеток в отторжении трансплантатов задокументирована как в клинике, так и в экспериментах на животных. Опосредуемое Т-клетками отторжение носит название клеточного отторжения, когда происходит разрушение клеток трансплантата цитотоксическими Т-лимфоцитами CD8+, и развиваются реакции гиперчувствительности замедленного типа, индуцированные активированными хелперными Т-клетками CD4+.
Основные антигенные различия между донором и реципиентом, приводящие к отторжению трансплантата, обусловлены различиями высокополиморфных аллелей HLA. Т-клетки реципиента распознают антигены трансплантата донора (аллогенные антигены, или аллоантигены) двумя путями:
- прямой путь распознавания состоит в том, что Т-клетки реципиента распознают аллогенные молекулы МНС донора на поверхности АПК, присутствующих в трансплантате. Считают, что дендритные клетки, переносимые реципиенту вместе с трансплантированным органом, представляют собой наиболее важные АПК, инициирующие ответ против трансплантата, т.к. они экспрессируют на высоком уровне не только молекулы МНС классов I и II, но и костимулирующие молекулы (например, В7-1 и В7-2).
Т-клетки организма-хозяина «встречают» дендритные клетки донора либо в трансплантированном органе, либо после миграции их в дренирующие лимфоузлы. Т-клетки CD8+ распознают молекулы МНС класса I и дифференцируются в активные цитотоксические Т-лимфоциты, способные уничтожать клетки трансплантата посредством описанных ранее механизмов. Хелперные Т-клетки CD4+ распознают аллогенные молекулы МНС класса II, пролиферируют и дифференцируются в эффекторные Тh1-клетки (а возможно, и Тh17-клетки).
Цитокины, секретируемые активированными Т-клетками CD4+, запускают реакцию гиперчувствительности замедленного типа в трансплантате, приводящую к повышению сосудистой проницаемости и местному накоплению мононуклеарных клеток (лимфоцитов и макрофагов), и трансплантат повреждается активированными макрофагами. Прямое распознавание аллогенных молекул МНС кажется парадоксальным, если вспомнить правило рестрикции по собственному МНС.
Если в обычных условиях распознавание Т-клеток ограничено чужеродными пептидами, презентированными собственными молекулами МНС, то почему эти клетки должны распознавать чужеродные молекулы МНС? Возможное объяснение состоит в том, что аллогенные молекулы МНС вместе со связанными пептидами имитируют комплексы собственных молекул МНС с пептидами, которые способны распознавать МНС-рестрикти-рованные Т-клетки. Таким образом, распознавание аллогенных молекул МНС представляет собой перекрестную реакцию Т-клеток, селеци-рованных для распознавания собственных молекул МНС и пептидов;
- в случае непрямого пути распознавания Т-лимфоциты реципиента распознают антигены МНС трансплантата донора после того, как они презентированы собственными АПК реципиента. Этот процесс включает захват и процессинг антигенов МНС и других чужеродных молекул трансплантата АПК реципиента.
Пептиды тканей донора презентируются собственными молекулами МНС реципиента, подобно любым другим чужеродным пептидам. Таким образом, непрямой путь сходен с физиологическим процессингом и презентацией других чужеродных антигенов (например, микробных). В непрямом пути образуются Т-клетки CD4+, проникающие в трансплантат и распознающие его антигены, презентированные АПК реципиента, также проникающими в трансплантат.
В результате развивается реакция гиперчувствительности замедленного типа. Однако цитотоксические Т-лимфоциты CD8+, которые могут возникнуть при непрямом пути, неспособны непосредственно распознавать и убивать клетки трансплантата, т.к. эти цитотоксические Т-лимфоциты распознают антигены трансплантата, презентированные АПК реципиента.
Следовательно, когда Т-клетки реагируют на трансплантат непрямым путем, основным механизмом клеточного отторжения может быть продукция Т-клеточных цитокинов и гиперчувствительность замедленного типа. Установлено, что прямой путь — основной при остром клеточном отторжении, тогда как непрямой путь наблюдается при хроническом отторжении. Однако такое разделение не является абсолютным.
б) Реакции, опосредуемые антителами. Хотя Т-клетки играют ведущую роль в отторжении трансплантированных органов, антитела, образующиеся к аллоантигенам трансплантатов, также служат важным фактором отторжения. Этот процесс называют гуморальным отторжением, он проявляется в двух формах. Сверхострое отторжение происходит в тех случаях, когда в кровотоке реципиента присутствуют преформированные антитела к антигенам донора. Такие антитела могут образоваться у реципиента после произошедшего ранее отторжения трансплантата почки.
Многорожавшие женщины, у которых образовались антитела анти-HLA к антигенам плода, унаследованным от отца, также имеют преформированные антитела к трансплантату, взятому от мужа либо детей или даже неродственных индивидов с аллелями HLA, общими с аллелями мужа. Предшествующая гемотрансфузия также может привести к пресенсибилизации, т.к. тромбоциты и лейкоциты богаты антигенами HLA, а донор и реципиент обычно не являются идентичными по HLA. При современной практике перекрестного тестирования, т.е. исследования сыворотки реципиента на присутствие антител к клеткам донора, сверхострое отторжение уже не является серьезной клинической проблемой.
У реципиентов, предварительно не сенсибилизированных к антигенам трансплантата, антигены HLA классов I и II донора способны вызвать продукцию антител. Эти антитела повреждают клетки донора разными механизмами, включая комплемент-зависимую цитотоксичность, воспаление и антитело-зависимую клеточную цитотоксичность. Первоначальной мишенью для этих антител при отторжении оказывается сосудистая система трансплантата. Таким образом, антитело-зависимое острое гуморальное отторжение обычно проявляется васкулитом, иногда называемым васкулитом отторжения.
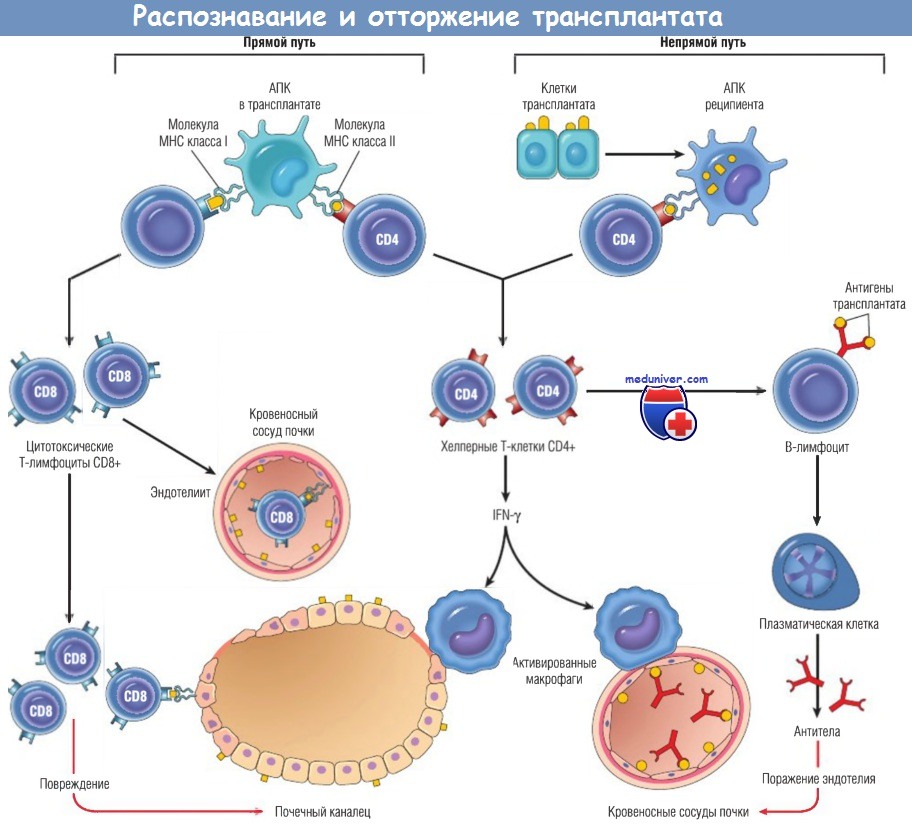
В случае прямого пути распознавания антигены МНС классов I и II донора на АПК в трансплантате
(вместе с костимуляторами, здесь не показанными) распознаются цитотоксическими Т-лимфоцитами CD8+ и хелперными Т-клетками CD4+ реципиента соответственно.
Клетки CD4+ пролиферируют и продуцируют цитокины (например, IFN-y), индуцирующие повреждения тканей в результате местной реакции гиперчувствительности замедленного типа.
Т-клетки CD8+, отвечающие на антигены трансплантата, дифференцируются в цитотоксические, которые убивают клетки трансплантата.
При непрямом пути распознавания антигены трансплантата захватываются, процессиру-ются и презентируются АПК реципиента, активирующими Т-клетки CD4+,
которые повреждают трансплантат в результате местной реакции гиперчувствительности замедленного типа и стимулируют продукцию антител В-лимфоцитами.
IFN — интерферон; МНС — главный комплекс гистосовместимости; АПК — антигенпрезентирующая клетка.
- Рекомендуем ознакомиться со следующей статьей "Механизмы отторжения трансплантата почки"
Оглавление темы "Болезни иммунной системы":- Причины и механизмы развития склеродермии (системного склероза)
- Морфология склеродермии (системного склероза)
- Клиника склеродермии (системного склероза)
- Механизмы отторжения трансплантатов органов
- Механизмы отторжения трансплантата почки
- Механизмы отторжения трансплантата костного мозга
- Первичные иммунодефициты и их характеристика
- Механизмы развития сцепленной с Х-хромосомой агаммаглобулинемии (агаммаглобулинемии Брутона)
- Механизмы развития общего вариабельного иммунодефицита
- Механизмы развития изолированного дефицита иммуноглобулина А (IgA)