
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Ген APC и его значение в развитии опухоли
Гены APC относятся к группе генов-супрессоров опухолей, основная функция которых заключается в снижении регуляторного влияния сигналов роста. Мутации APC в половых клетках (5q21) характерны для семейного аденоматозного полипоза, при котором у индивидов, рожденных с одним мутантным аллелем, появляются сотни и тысячи аденоматозных полипов в толстой кишке в подростковом возрасте или до 30 лет. Почти всегда один полип или несколько подвергаются злокачественной трансформации после накопления мутаций в клетках полипов.
Подобно ситуациям с другими генами-супрессорами опухолей, для формирования неоплазии должны отсутствовать обе копии гена APC. Данное заключение было подтверждено в экспериментальном аденоматозном полипозе на мышах, индуцированном таргетной деструкцией APC в слизистой толстой кишки. Как будет показано далее, для маглинизации опухоли из аденоматозного полипа необходимо накопление еще нескольких мутаций. Помимо семейных случаев рака толстой кишки гомозиготная утрата генов APC выявляется в 70-80% колоректальных карцином и спорадических аденом, что подтверждает участие APC в патогенезе опухолей толстой кишки.
APC является компонентом сигнального пути WNT. Основная роль APC заключается в контроле метаболизма клеток, адгезии и полярности в ходе эмбриогенеза. Сигнальный путь WNT используется кроветворными стволовыми клетками при их обновлении. Сигнальный путь WNT содержит несколько поверхностных рецепторов, называемых FRZ-рецепторами, стимулирующих b-катенин и APC.
Важнейшая функция белка APC — подавление b-катенина. При нефункционирующем сигнальном пути WNT APC разрушает b-катенин, препятствуя его накоплению в цитоплазме. При этом формируется макромолекулярный комплекс из b-катенина, аксина и GSK3b, вызывающий фосфорилирование b-катенина и связывание его с убиквитином, а впоследствии и деструкцию в протеасомах.
Сигнальный путь WNT дезактивирует деструктивный комплекс APC-аксин GSK3P, что обеспечивает трансляцию b-катенина в ядро клетки. В ядре клетки b-катенин формирует комплекс с TGF, трансформирующим фактором роста, индуцирующим клеточную пролиферацию через усиление транскрипции с-MYC, циклина D1 и других генов. При инактивации гена APC и деструктивного комплекса b-катенин сохраняется, перемещается в ядро, кооперируется с TGF и активирует транскрипцию.
Таким образом, клетки с утратой APC ведут себя так, будто они находятся под постоянным влиянием сигнального пути WNT. Значимость пути APC/ b-катенина для онкогенеза подтверждается наблюдением пациента, больного раком толстой кишки, имеющего нормальный ген APC и мутантный b-катенин, что исключает его деструкцию под действием APC и приводит к накоплению мутантного b-катенина в ядре клетки.
Дисрегуляция пути APC/b-катенина наблюдается не только при раке толстой кишки; мутации гена b-катенина наблюдаются более чем в 50% нейробластом и примерно в 20% гепатоцеллюлярных карцином.
b-катенин связывается с цитоплазматической частью Е-кадгерина, поверхностного белка клетки, участвующего в межклеточной адгезии. Утрата межклеточных контактов, происходящая в ранах и при повреждении эпителия, приводит к разобщению р-катенина и Е-кадгерина, что позволяет b-катенину мигрировать в ядро и стимулировать пролиферацию клетки. Такая последовательность событий наблюдается при адекватной реакции клетки на повреждение, что позволяет организму репарировать очаги повреждения и раны. Восстановление связывания с Е-кадгерином в заживших ранах приводит к секвестрации в мембране b-катенина и редукции пролиферативного сигнала. Об этих процессах говорят, что клетки подвергаются контактному торможению.
Отсутствие контактного торможения вследствие нарушений связи Е-кадгерина с b-катенином или других процессов является ключевой характеристикой карцином. Более того, отсутствие кадгерина может способствовать формированию злокачественного фенотипа за счет дезагрегации опухолевых клеток, способствующей локальному инвазивному росту и метастазиро-ванию. Снижение экспрессии Е-кадгерина на поверхности опухолевых клеток происходит во многих видах злокачественных опухолей: пищевода, толстой кишки, молочной железы и предстательной железы. Мутации Е-кадгерина в половых клетках повышают предрасположенность к семейной карциноме желудка.
Кроме того, мутация и снижение экспрессии Е-кадгерина отмечаются при диффузном раке желудка. Молекулярные механизмы снижения экспрессии Е-кадгерина могут быть различными. В небольшом количестве наблюдений злокачественных опухолей обнаруживают мутации гена Е-кадгерина (локализованного на 16q), в других случаях снижение экспрессии Е-кадгерина происходит вторично в результате мутаций генов b-катенина. Также Е-кадгерин может быть блокирован репрессорами транскрипции, например SNAIL, что наблюдается при эпителиально-мезенхимальной трансдифференцировке и метастазировании.
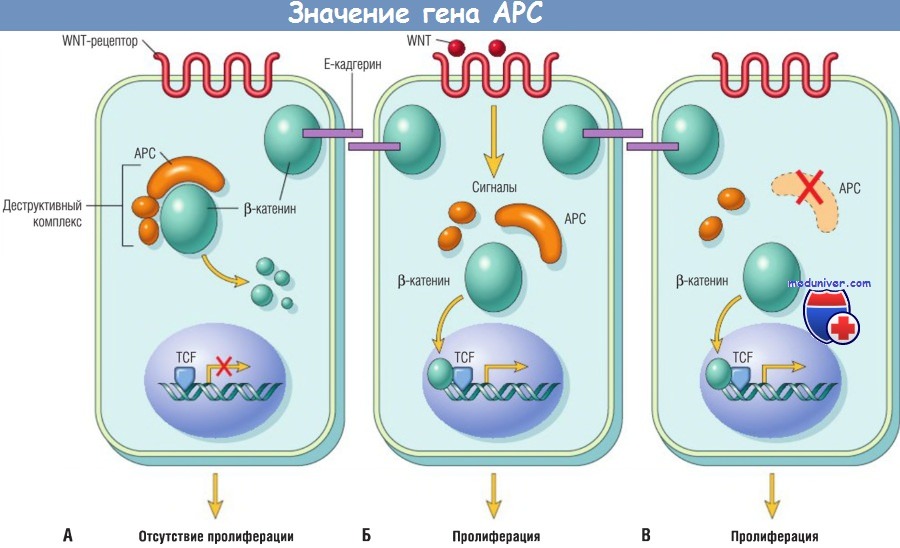
В покоящихся клетках (с нефункционирующим WNT) р-катенин формирует макромолекулярный комплекс с белком АРС. В этом комплексе b-катенин разрушается, в результате внутриклеточный уровень b-катенина снижается.
(Б) При стимуляции клетки молекулами WNT деструктивный комплекс дезактивируется, деградации b-катенина не происходит, в результате увеличивается его уровень в цитоплазме, b-катенин перемещается в ядро, где связывается с TCF, фактором транскрипции, который активирует несколько генов, вовлеченных в клеточный цикл.
(В) При мутации или отсутствии гена АРС разрушение b-катенина невозможно, b-катенин перемещается в ядро и коактивирует гены, запускающие клеточный цикл, при этом клетки ведут себя, как будто они находятся под постоянной стимуляцией сигнального пути WNT.
TCF — трансформирующий фактор роста.
- Рекомендуем ознакомиться со следующей статьей "Ген INK4a/AKF (локус CDKN2A) и его значение в развитии опухоли"
Оглавление темы "Патофизиология онкологических заболеваний":- Онкоген RAS и его характеристика
- Изменения нерецепторных тирозинкиназ при развитии рака
- Протоонкоген MYC в развитии опухолей
- Циклины и циклин-зависимые киназы в развитии опухолей
- Гены-супрессоры опухоли и их характеристика
- Ген RB и его значение в развитии опухоли
- Ген p53 и его значение в развитии опухоли
- Ген APC и его значение в развитии опухоли
- Ген INK4a/AKF (локус CDKN2A) и его значение в развитии опухоли
- Сигнальный путь TGF-b и его значение в развитии опухоли