
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Дифференцировка прогениторных клеток. Стволовые свойства культурных клеток
В последние годы неоднократно указывалось или предполагалось, что прогениторные клетки в целом ряде различных постнатальных тканей имеют очевидный неортодоксальный потенциал дифференциации. В частности, показано, что костный мозг содержит системно мигрирующие миогенные клетки-предшественники (Ferrari et al, 1998), нейральные стволовые клетки способны рекапитулировать гемопоэз у облученных мышей (Mezey et al., 2000), клетки костного мозга генерируют нейральные прогениторы (Mezey et al., 2000) и гепатоциты (Lagasse et al., 2000), а стромальные клетки костного, мозга обладают нейрогенным потенциалом (Кореи et al., 1999; Woodbury et al., 2000). Поражает эволюционно разделенная природа клеток-предшественников и терминальных клеток.
Дифференциация в пределах производных разных зародышевых слоев нарушает общий закон эволюционной биологии. Хотя общие законы не являются догмами (в частности, зародышевые слои более важны для эмбриологов, чем для самого эмбриона), по-прежнему остается неоспоримым тот факт, что даже в эмбрионе клетки с трансгермальным потенциалом существуют только в определенных строгих пространственно-временных параметрах (за исключением клеток нервного гребня, которые, несмотря на нейроэктодермальное происхождение, генерируют черепно-лицевые "мезодермальные" ткани, включая кости). Клетки, выращенные в культуре из внутренней клеточной массы, самообновляются и сохраняют плюрипотентность в культуре в течение длительного времени. Однако это своего рода артефакт, о котором нам известны лишь некоторые "почему" и "для чего" (фидерный слой клеток, LIF).
В процессе эмбриогенеза эмбриональные стволовые клетки способны сохранять плюрипотентность и свойство самообновления очень недолго, после чего переходят в мульти- и полипотентные клетки коммитированных зародышевых линий (Bianco et al., 2001). Возникает ключевой вопрос о происхождении мультипотентных клеток постнатальных организмов. Если стромальные клетки костного мозга входят в состав диффузной системы постнатальных мультипотентных стволовых клеток и в то же время имеют васкуло-перицитное происхождение, тогда логично допустить, что микрососудистая система представляет собой резервуар мультипотентных клеток во многих, если не во всех тканях (Bianco, Cossu, 1999).
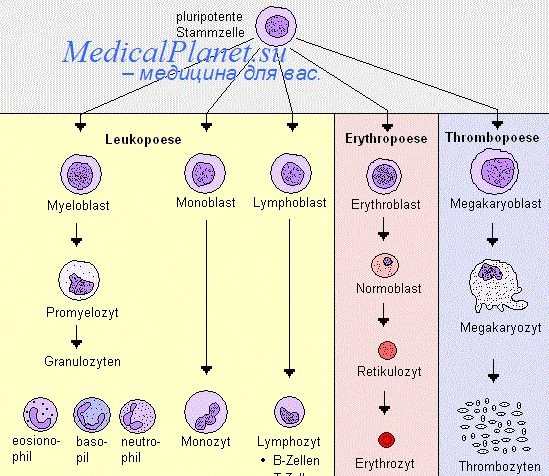
Однако если мультипотентные клетки в зрелом организме диффузно распространены повсеместно, то каким образом блокируется тератомогенез? К тому же дифференцированные клетки зрелых тканей должны обладать сигнальными механизмами не только блокировки образования тератом, но и предупреждения формирования эктопических тканей, что необходимо для сохранения самоидентичности. Третий проблемный вопрос касается того, насколько "стволовые свойства" (самообновление и мультипотентность) экспериментальных клеточных систем определяются манипуляциями на клетках in vitro? Действительно ли in vivo имеются клетки с подобными характеристиками, или эти неизвестные и неожиданные эффекты всего лишь следствия манипулирования клетками в культуре?
В частности, до какой степени условия культивирования клеток имитируют эффекты энуклеированной ооцитной цитоплазмы, которая позволяет ядру соматической клетки генерировать организм, подобный клонированной овце Долли? Новое, биотехнологически корректное определение того, что представляет собой стволовая клетка, должно включать условия, при которых происходит фиксация феномена, а не догадки на основании сведений, полученных ex vivo, об истинных свойствах объекта in vivo. Даже для праматери всех стволовых клеток — ЭСК — самообновление и плюрипотентность ограничены определенными сроками и событиями in vivo, но менее ограничены ex vivo. Справедливо ли это по отношению к другим стволовым клеткам? Стромальные стволовые клетки костного мозга, например, могут экстенсивно размножаться в культуре, но большинство из них никогда не делятся in vivo после прекращения роста скелета (за исключением нескольких клеток, которые участвуют в ремоделировании костной ткани и в реакциях на травму или заболевание).
Какой физиологический механизм отвечает за активацию стволовых ресурсов in vivo? Все эти вопросы имеют огромное значение не только с философской или эзотерической, но и с практической точки зрения. Ответы хотя бы на некоторые их них уже позволят начать разработку адекватных биотехнологических методов клинического применения уникальных свойств стволовых клеток в медицине (Bianco et al., 2001).
- Читать далее "Определение потенциала дифференцировки стволовых клеток. Системная трансплантация стволовых клеток"
Оглавление темы "Нейральные стволовые клетки":1. Дифференцировка прогениторных клеток. Стволовые свойства культурных клеток
2. Определение потенциала дифференцировки стволовых клеток. Системная трансплантация стволовых клеток
3. Доставка стволовых клеток в ткани. Генная инженерия клеток
4. Лечение рецессивных заболеваний. Преобразованные стволовые клетки
5. Коррекция генетического заболевания. Нейтральные стволовые клетки
6. Стволовой резерв нервной системы. Нейральные стволовые клетки эмбриона
7. Маркеры нейральных стволовых клеток. Дифференциация стволовых клеток нервной системы
8. Регенерация центральной нервной системы. Трансплантация нейральных стволовых клеток
9. Культура нейральных стволовых клеток. Приживление нейральных стволовых клеток у разных видов
10. Ксенотрансплантация нейральных стволовых клеток. Получение нейральных стволовых клеток