
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Миогенная дифференцировка стволовых клеток. Факторы дифференцировки стволовых клеток
Трансфекция в ЭСК конструкции, собранной из гена аминогликозидфосфотрансферазы под промотором тяжелой цепи миозина, позволяет получить мигрирующую популяцию кардиомиоцитов, способных активно встраиваться в мышечный синцитий реципиента (Leiden, 1999; Rittenger, 1999).
При нормальном развитии зародыша начальные этапы миогенной дифференцировки клеток in vivo инициируются индукцией синтеза транскрипционных факторов. Период раннего миогенеза контролируется тканеспецифическими регуляторами семейства bHLH, которые активируют транскрипцию мышечных генов. В ранних миобластах экспрессируются гены myoD и ту/5, а на более поздних этапах миогенеза в миотубах и миофибриллах появляются myogenin и mrf4 (Olson, 1992; Olson, Klein, 1994; Озернюк, 1998).
В культуре ЭСК первые миобласты возникают на 4-е (линия BLC6) или 5-7-е сутки (линия D3) после прикрепления эмбриоидных телец к желатиновой подложке (Rohwedel et al., 1994; Guan et al., 1999). Затем, через 24-48 часов, миобласты сливаются в миотубы. Первыми экспрессируются гены семейства bHLH, прежде всего ту/5. Несколько позднее наблюдается экспрессия генов myogenin и myoD, а также гена М-кадгерина, кодирующего структурный белок М-линии. На терминальных стадиях дифференцировки в миотубах экспрессируется ген ту/6.
Наиболее ранняя экспрессия показана для тинина и небулина, несколько позднее синтезируются М-кадгерин, миогенин и тяжелые цепи саркомерного миозина. Таким образом, последовательность экспрессии генов и синтеза белков в процессе дифференцировки мышечной ткани in vitro в целом подобна таковой in vivo (Мануйлова и др., 2001).
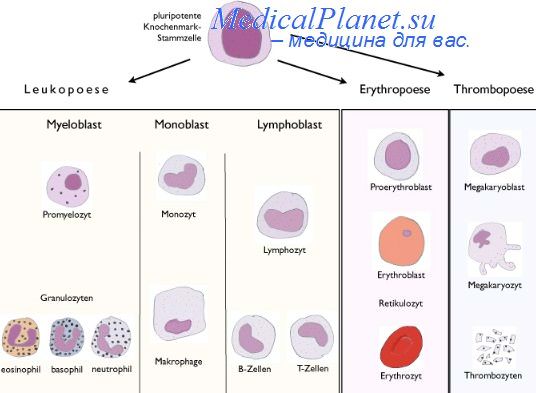
В настоящее время наиболее изучены дифференцировочные эффекты таких факторов роста как FGF, TGF, активин-А, ВМР-4, HGF, EGF, NGF и ретиновая кислота (Shuldiner, 2000). По дифференцирующему эффекту, выявленному в ряде исследований, данные факторы разделяют на три категории. В первую группу включены факторы роста, индуцирующие, главным образом, формирование мезодермальных клеток (активин А и TGF).
Во вторую вошли факторы, активирующие экспрессию экто- и мезодермальных маркеров (ретиноевая кислота, EGF, ВМР-4 и FGF). Третья категория представлена факторами, которые позволяют индуцировать дифференцировку ЭСК в клетки всех трех эмбриональных зародышевых листков (Odorico et al., 2001). В последнее время появились сообщения о дифференцирующем действии аскорбиновой кислоты, индуцирующей образование из ЭСК человека кардиомиоцитов (Takahashi et al., 2003).
Перед анализом терапевтической эффективности трансплантации ЭСК и их производных резюмируем приведенный выше материал. Возможности ЭСК в плане полной реализации эмбриогенеза in vitro недостаточны, поскольку дефекты развития в этом случае обусловлены отсутствием мезенхимальных стволовых клеток, которые в организме возникают автономно и независимо от ЭСК.
Генетические потенции ЭСК меньше генетического потенциала зиготы, поэтому непосредственно для клонирования зародышей ЭСК не используются. Уникальный биологический потенциал ЭСК как единственных клеток, в которых программы развития развернуты в полном объеме последовательной реализации, находит применение в исследованиях по изучению функции генов. С помощью ЭСК проводится расшифровка первых комбинаций сигналов, активирующих экспрессию ранних и поздних генов, кодирующих развитие трех зародышевых листков.
Сохранение плюрипотентности генома ЭСК in vitro делает их уникальным инструментом для репаративной регенерации, способным в автоматическом режиме восполнять клеточные потери при повреждении органов и тканей. В идеальном гипотетическом варианте можно допустить, что "... при трансплантации донорских ЭСК в организм реципиента переносятся компактно упакованные программы, которые при благоприятных условиях реализуются в строительство новой ткани", способной "... эффективно встраиваться в организм реципиента как на морфологическом, так и функциональном уровне" (Репин, 2001).
- Читать далее "Активность дифференцированных стволовых клеток. Пересадка эмбриональных стволовых клеток"
Оглавление темы "Дифференциация и применение стволовых клеток":1. Программы органогенеза из стволовых клеток. Формирование зародышевых листков
2. Развитие нервной ткани из стволовых клеток. Формирование кроветворения из стволовых клеток
3. Развитие клеток крови из стволовых клеток. Формирование кардиомиоцитов из стволовых клеток
4. Миогенная дифференцировка стволовых клеток. Факторы дифференцировки стволовых клеток
5. Активность дифференцированных стволовых клеток. Пересадка эмбриональных стволовых клеток
6. Применение эмбриональных стволовых клеток. Эмбриональные стволовые клетки при иммунодефицитах
7. Трансплантация нейробластов. Нейротрансплантация в лечении паркинсонизма
8. Клинический потенциал стволовых клеток. Тератомы эмбриональных стволовых клеток
9. Терапевтичекое клонирование стволовых клеток. Принципы терапевтического клонирования
10. Размножение стволовых клеток. Регенеративно-пластические технологии и биоэтические вопросы