
MedicalPlanet
Контактные данные:
admin@medicalplanet.su
2009-2023
Развитие клеток крови из стволовых клеток. Формирование кардиомиоцитов из стволовых клеток
Клетки желточного мешка переводят в гемопоэтические линии путем добавления в среду культивирования IL-3 и супернатанта фидера от стромаль-ных клеток. Полагают, что фидер продуцирует целый комплекс сигналов для созревания линий кроветворных клеток: IL-3, SCF, FGF, IGF1, IL-6, G-CSF (Palacious, 1995).
Трансфекция в геном ЭСК гена hox-b4 значительно повышает скорость образования предшественников миелоидного и эритроидного ряда in vitro. Трансфицированные геноhox-b4 ЭСК обладают 50-кратно усиленным потенциалом кроветворения за счет резкого увеличения численности ранних прогениторных популяций гемопоэтических стволовых клеток.
Кроме того, трансплантация таких искусственно созданных кроветворных клонов с измененной пропорцией стволовых/прогениторных клеток оказалась значительно более эффективной в плане колонизации, приживления в костном мозге реципиента и выработки форменных элементов крови (Hegalson, 1996; Robey, 2000).
Порядок экспрессии генов и белков в ходе образования из ЭСК кардиомиоцитов также напоминает последовательность этих процессов in vivo. При дифференцировке кардиомиоцитов из ЭСК первые сокращения групп клеток отмечены через 1-2 суток после прикрепления эмбриоидных телец к субстрату, а максимальное их число — через 5-7 суток (Wobus et aL, 1991; Guan et aL, 1999).
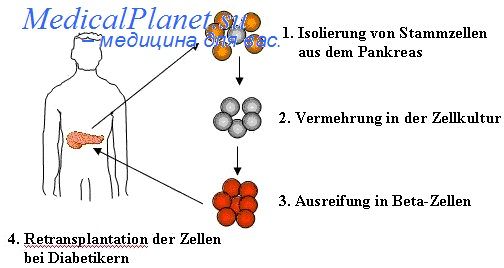
Первыми в ходе кардиогенеза экспрессируются гены специфических транскрипционных факторов Nkx 2,5. В клетках линии D3 это происходит на 3-й сутки (Wobus, Guan, 1998). Однако на еще более ранних этапах (за двое суток до начала сокращений кардиомиоцитов) наблюдается экспрессия гена, кодирующего субъединицу белка кальциевого канала. Затем следует экспрессия генов а и Р тяжелых цепей миозина сердца, а на более поздних стадиях дифференцировки активируются ген легких цепей миозина 2v (mlc-2v) и ген атриального натрийуретического фактора.
Последовательность экспрессии белков в ходе кардиогенеза выглядит следующим образом. Первыми экспрессируются тинин (специфический белок Z-линии), а-актинин и миомезин, затем саркомерные тяжелые цепи миозина и а-актина, а еще позднее — М-белок (Wobus, Guan, 1998).
Сейчас разработаны более эффективные методы получения кардиомиоцитов под контролем экспрессии гена crypto. Продуктом этого гена является фактор транскрипции, контролирующий первый этап генетической детерминации кардиомиоцитов. Имплантация в бластоцисту клеток линии ЭСК с двойным нокаутом (crypto) приводит у химерных зародышей к потере способности генерировать зачаток сердца и кардиомиоциты.
Установлено, что накопление кислородных радикалов и электростимуляция ЭСК в культуре способствуют образованию зрелых сокращающихся кардиомиоцитов с завершенной структурой саркомеров. Полученные таким методом кардиомиоциты хорошо встраиваются в миокард развивающихся зародышей или в постишемическую некробрютическую ткань сердца (Репин, 2001).
- Читать далее "Миогенная дифференцировка стволовых клеток. Факторы дифференцировки стволовых клеток"
Оглавление темы "Дифференциация и применение стволовых клеток":1. Программы органогенеза из стволовых клеток. Формирование зародышевых листков
2. Развитие нервной ткани из стволовых клеток. Формирование кроветворения из стволовых клеток
3. Развитие клеток крови из стволовых клеток. Формирование кардиомиоцитов из стволовых клеток
4. Миогенная дифференцировка стволовых клеток. Факторы дифференцировки стволовых клеток
5. Активность дифференцированных стволовых клеток. Пересадка эмбриональных стволовых клеток
6. Применение эмбриональных стволовых клеток. Эмбриональные стволовые клетки при иммунодефицитах
7. Трансплантация нейробластов. Нейротрансплантация в лечении паркинсонизма
8. Клинический потенциал стволовых клеток. Тератомы эмбриональных стволовых клеток
9. Терапевтичекое клонирование стволовых клеток. Принципы терапевтического клонирования
10. Размножение стволовых клеток. Регенеративно-пластические технологии и биоэтические вопросы